
В прокариотах . Комплементарная ДНК также образуется ретровирусами (ВИЧ-1 , ВИЧ-2 , Вирусом иммунодефицита обезьян) и затем интегрируется в ДНК хозяина, образуя провирус .
Часто гены эукариот удается экспрессировать в клетках прокариот. В наиболее простом случае, метод предполагает встраивание эукариотической ДНК в геном прокариот, далее транскрипцию ДНК в мРНК и затем трансляцию мРНК в белки. Клетки прокариот не имеют ферментов для вырезания интронов, и поэтому интроны из ДНК эукариот должны быть вырезаны до момента встраивания в геном прокариот. ДНК, комплементарная зрелой мРНК, таким образом, называется комплементарной ДНК — cDNA (кДНК). Для успешной экспрессии белков, закодированных в эукариотической cDNA в прокариотах, требуются также регуляторные элементы прокариотических генов (например, промоторы).
Одним из методов для получения необходимого гена (молекулы ДНК), которая будет подлежать репликации (клонированию) с выходом значительного количества реплик, является конструирование на мРНК комплементарной относительно неё ДНК (кДНК). Этот метод требует применения обратной транскриптазы — фермента, который присутствует в некоторых РНК-содержимых вирусах и обеспечивает синтез ДНК на РНК матрице.
Метод широко применяется для получения кДНК и включает в себя выделение из тотальной мРНК ткани мРНК, которая кодирует трансляцию определенного белка (например, интерферона, инсулина) с дальнейшим синтезом на этой мРНК как на матрице необходимой кДНК с помощью обратной транскриптазы.
Ген, который был получен с помощью вышеуказанной процедуры (кДНК), необходимо ввести в бактериальную клетку таким образом, чтобы он интегрировался в её геном. Для этого формируют рекомбинантную ДНК, которая состоит из кДНК и особенной молекулы ДНК, которая правит как проводник, или вектор, способный проникать реципиенту в клетку. В роли векторов для кДНК применяют вирусы или плазмиды. Плазмиды — это небольшие кольцевые молекулы ДНК, которые находятся отдельно от нуклеоида бактериальной клетки, содержат в своем составе несколько важных для функции всей клетки генов (например, гены стойкости к антибиотикам и могут реплицироваться независимо от основного генома (ДНК) клетки. Биологически важными и практически полезными для генной инженерии свойствами плазмида являются их способность к переходу из одной клетки в другую по механизму трансформации или конъюгации, а также способность включаться в бактериальную хромосому и реплицироваться вместе с ней.
Напишите отзыв о статье «Комплементарная ДНК»
Примечания
Просмотр этого шаблонаАзотистые основанияНуклеозидыНуклеотидыРНК 11pxДНК 11pxАналогиТипы векторов
Отрывок, характеризующий Комплементарная ДНК
– А эти «ключики», они разве никогда не повторяются у других? – решила продолжить свои расспросы я. – Нет, но иногда бывает кое-что другое…– почему-то забавно улыбаясь, ответила крошка. – Я в начале именно так и попалась, за что меня очень даже сильно «потрепали»… Ой, это было так глупо!.. – А как? – очень заинтересовавшись, спросила я. Стелла тут же весело ответила: – О, это было очень смешно! – и чуть подумав, добавила, – но и опасно тоже… Я искала по всем «этажам» прошлое воплощение своей бабушки, а вместо неё по её «ниточке» пришла совсем другая сущность, которая как-то сумела «скопировать» бабушкин «цветок» (видимо тоже «ключик»!) и, как только я успела обрадоваться, что наконец-то её нашла, эта незнакомая сущность меня безжалостно ударила в грудь. Да так сильно, что у меня чуть душа не улетела!.. – А как же ты от неё избавилась? – удивилась я. – Ну, если честно, я и не избавлялась… – смутилась девочка. – Я просто бабушку позвала… – А, что ты называешь «этажами»? – всё ещё не могла успокоиться я. – Ну, это разные «миры» где обитают сущности умерших… В самом красивом и высоком живут те, которые были хорошими… и, наверное, самыми сильными тоже. – Такие, как ты? – улыбнувшись, спросила я. – О, нет, конечно! Я наверное сюда по ошибке попала. – Совершенно искренне сказала девчушка. – А знаешь, что самое интересное? Из этого «этажа» мы можем ходить везде, а из других никто не может попасть сюда… Правда – интересно?.. Да, это было очень странно и очень захватывающе интересно для моего «изголодавшегося» мозга, и мне так хотелось узнать побольше!.. Может быть потому, что до этого дня мне никогда и никто ничего толком не объяснял, а просто иногда кто-то что-то давал (как например, мои «звёздные друзья»), и поэтому, даже такое, простое детское объяснение уже делало меня необычайно счастливой и заставляло ещё яростнее копаться в своих экспериментах, выводах и ошибках… как обычно, находя во всём происходящем ещё больше непонятного. Моя проблема была в том, что делать или создавать «необычное» я могла очень легко, но вся беда была в том, что я хотела ещё и понимать, как я это всё создаю… А именно это пока мне не очень-то удавалось… – А остальные «этажи»? Ты знаешь, сколько их? Они совсем другие, непохожи на этот?.. – не в состоянии остановиться, я с нетерпением заваливала Стеллу вопросами. – Ой, я тебе обещаю, мы обязательно пойдём туда погулять! Ты увидишь, как там интересно!.. Только там и опасно тоже, особенно в одном. Там такие чудища гуляют!.. Да и люди не очень приятные тоже. – Я думаю, я уже видела похожих чудищ, – кое-что вспомнив, не очень уверенно сказала я. – Вот посмотри… И я попробовала показать ей первых, встреченных в моей жизни, астральных существ, которые нападали на пьяного папу малышки Весты. – Ой, так это же такие же! А где ты их видела? На Земле?!.. – Ну, да, они пришли, когда я помогала одной хорошей маленькой девочке проститься со своим папой… – Значит, они приходят и к живым?.. – очень удивилась моя подружка. – Не знаю, Стелла. Я ещё вообще почти ничего не знаю… А так хотелось бы не ходить в потёмках и не узнавать всё только на «ощупь»… или из своего опыта, когда постоянно за это «бьют по голове»… Как ты думаешь, твоя бабушка не научила бы чему-то и меня?.. – Не знаю… Ты, наверное, должна сама у неё об этом спросить? Девочка глубоко о чём-то задумалась, потом звонко рассмеялась и весело сказала: – Это было так смешно, когда я только начала «творить»!!! Ой, ты бы знала, как это было смешно и забавно!.. Вначале, когда от меня «ушли» все, было очень грустно, и я много плакала… Я тогда ещё не знала где они, и мама, и братик… Я не знала ещё ничего. Вот тогда, видимо, бабушке стало меня жалко и она начала понемножку меня учить. И… ой, что было!.. Вначале я куда-то постоянно проваливалась, создавала всё «шиворот навыворот» и бабушке приходилось за мной почти всё время наблюдать. А потом я научилась… Даже жалко, потому что она теперь уже реже приходит… и я боюсь, что может когда-нибудь она не придёт совсем…
В реакции, катализируемой обратной транскриптазой .
кДНК часто используется для клонирования генов эукариот в прокариотах . Комплементарная ДНК также образуется ретровирусами (ВИЧ-1 , ВИЧ-2 , Вирусом иммунодефицита обезьян) и затем интегрируется в ДНК хозяина, образуя провирус .
Часто гены эукариот удается экспрессировать в клетках прокариот. В наиболее простом случае, метод предполагает встраивание эукариотической ДНК в геном прокариот, далее транскрипцию ДНК в мРНК и затем трансляцию мРНК в белки. Клетки прокариот не имеют ферментов для вырезания интронов, и поэтому интроны из ДНК эукариот должны быть вырезаны до момента встраивания в геном прокариот. ДНК, комплементарная зрелой мРНК, таким образом, называется комплементарной ДНК — cDNA (кДНК). Для успешной экспрессии белков, закодированных в эукариотической cDNA в прокариотах, требуются также регуляторные элементы прокариотических генов (например, промоторы).
Одним из методов для получения необходимого гена (молекулы ДНК), которая будет подлежать репликации (клонированию) с выходом значительного количества реплик, является конструирование на мРНК комплементарной относительно неё ДНК (кДНК). Этот метод требует применения обратной транскриптазы — фермента, который присутствует в некоторых РНК-содержимых вирусах и обеспечивает синтез ДНК на РНК матрице.
Метод широко применяется для получения кДНК и включает в себя выделение из тотальной мРНК ткани мРНК, которая кодирует трансляцию определенного белка (например, интерферона, инсулина) с дальнейшим синтезом на этой мРНК как на матрице необходимой кДНК с помощью обратной транскриптазы.
Ген, который был получен с помощью вышеуказанной процедуры (кДНК), необходимо ввести в бактериальную клетку таким образом, чтобы он интегрировался в её геном. Для этого формируют рекомбинантную ДНК, которая состоит из кДНК и особенной молекулы ДНК, которая правит как проводник, или вектор, способный проникать реципиенту в клетку. В роли векторов для кДНК применяют вирусы или плазмиды. Плазмиды — это небольшие кольцевые молекулы ДНК, которые находятся отдельно от нуклеоида бактериальной клетки, содержат в своем составе несколько важных для функции всей клетки генов (например, гены стойкости к антибиотикам и могут реплицироваться независимо от основного генома (ДНК) клетки. Биологически важными и практически полезными для генной инженерии свойствами плазмида являются их способность к переходу из одной клетки в другую по механизму трансформации или конъюгации, а также способность включаться в бактериальную хромосому и реплицироваться вместе с ней.
Репликация– удвоение молекул ДНК, в результате которого образуются две двойные спирали ДНК. Основано на принципах:
1) Комплементарность – каждая из двух цепей – матрица для синтеза комплементарной цепи. Это свойство обеспечивается особенностями химической организации молекулы ДНК, состоящей из двух комплементарных цепей. В процессе репликации на каждой полинуклеотидной цепи материнской молекулы ДНК синтезируется комплементарная ей цепь. В итоге из одной двойной спирали ДНК образуются две идентичные двойные спирали.
2) Полуконсервативность – каждая из двух двойных спиралей несет одну нить
материнской ДНК.
3) Антипараллельность– каждая цепь ДНК имеет определенную ориентацию: 5″-конец одной цепи соединяется с 3″-концом другой, и наоборот.
4) Прерывистость – репликация осуществляется фрагментами.
Инициация репликации осуществляется в особых участках ДНК, обозначаемых ori(от англ. origin -начало). Они включают последовательность, состоящую из 300 нуклеотидных пар, узнаваемую специфическими белками. Двойная спираль ДНК в этих локусах разделяется на две цепи, при этом, как правило, по обе стороны от точки начала репликации образуются области расхождения полинуклеотидных цепей — репликационные вилки, которые движутся в противоположных от локуса ori направлениях. Между репликационными вилками образуется структура, называемая репликационным глазком, где на двух цепях материнской ДНК образуются новые полинуклеотидные цепи.
С помощью фермента хеликазы, разрывающего водородные связи, двойная спираль ДНК расплетается в точках начала репликации. Образующиеся при этом одинарные цепи ДНК связываются специальными дестабилизирующими белками, которые растягивают остовы цепей, делая их азотистые основания доступными для связывания с комплементарными нуклеотидами, находящимися в нуклеоплазме. На каждой из цепей, образующихся в области репликационной вилки, при участии фермента ДНК-полимеразы осуществляется синтез комплементарных цепей.
В процессе синтеза репликационные вилки движутся вдоль материнской спирали в противоположных направлениях, захватывая все новые зоны.
Разделение спирально закрученных цепей родительской ДНК ферментом хеликазой вызывает появление супервитков перед репликационной вилкой. Это объясняется тем, что при расхождении каждых 10 пар нуклеотидов, образующих один виток спирали, родительская ДНК должна совершить один полный оборот вокруг своей оси. Следовательно, для продвижения репликационной вилки вся молекула ДНК перед ней должна была бы быстро вращаться, что потребовало бы большой затраты энергии. В действительности это не наблюдается благодаря особому классу белков, называемых ДНК-топоизомеразами. Топоизомераза разрывает одну из цепей ДНК, что дает ей возможность вращаться вокруг второй цепи. Это ослабляет накопившееся напряжение в двойной спирали ДНК.
К высвобождающимся водородным связям нуклеотидных последовательностей разделенных родительских цепей присоединяются свободные нуклеотиды из нуклеоплазмы, где они присутствуют в виде дезоксирибонуклеозидгрифосфатов: дАТФ, дГТФ, дЦТФ, дТТФ. Комплементарный нуклеозидтрифосфат образует водородные связи с определенным основанием материнской цепи ДНК. Затем при участии фермента ДНК-полимеразы он связывается фосфодиэфирной связью с предшествующим нуклеотидом вновь синтезируемой цепи, отдавая при этом неорганический пирофосфат.
Поскольку ДНК-полимераза присоединяет очередной нуклеотид к ОН-группе в 3″-положении предшествующего нуклеотида, цепь постепенно удлиняется на ее 3″-конце.
Особенностью ДНК-полимеразы является ее неспособность начать синтез новой полинуклеотидной цепи путем простого связывания двух нуклеозидтрифосфатов: необходим 3″-ОН-конец какой-либо полинуклеотидной цепи, спаренной с матричной цепью ДНК, к которой ДНК-полимераза может лишь добавлять новые нуклеотиды. Такую полинуклеотидную цепь называют затравкойили праймером.
Роль затравки для синтеза полинуклеотидных цепей ДНК в ходе репликации выполняют короткие последовательности РНК, образуемые при участии фермента РНК-праймазы. Указанная особенность ДНК-полимеразы означает, что матрицей при репликации может служить лишь цепь ДНК, несущая спаренную с ней затравку, которая имеет свободный 3″-ОН-конец.
Способность ДНК-полимеразы осуществлять сборку полинуклеотида в направлении от 5″- к 3″ -концу при антипараллельном соединении двух цепей ДНК означает, что процесс репликации должен протекать на них по-разному. Действительно, если на одной из матриц (3″ → 5″) сборка новой цепи происходит непрерывно от 5″- к 3″-концу и она постепенно удлиняется на 3″-конце, то другая цепь, синтезируемая на матрице (5″ → 3″), должна была бы расти от 3″- к 5″-концу. Это противоречит направлению действия фермента ДНК-полимеразы.
В настоящее время установлено, что синтез второй цепи ДНК осуществляется короткими фрагментами (фрагменты Оказаки) также в направлении от 5″- к 3″-концу (по типу шитья «назад иголкой»). У прокариот фрагменты Оказаки содержат от 1000 до 2000 нуклеотидов, у эукариот они значительно короче (от 100 до 200 нуклеотидов). Синтезу каждого такого фрагмента предшествует образование РНК-затравки длиной около 10 нуклеотидов. Вновь образованный фрагмент с помощью фермента ДНК-лигазы соединяется с предшествующим фрагментом после удаления его РНК-затравки.
В связи с указанными особенностями репликационная вилка является асимметричной. Из двух синтезируемых дочерних цепей одна строится непрерывно, ее синтез идет быстрее и эту цепь называют лидирующей. Синтез другой цепи идет медленнее, так как она собирается из отдельных фрагментов,требующих образования, а затем удаления РНК-затравки. Поэтому такую цепь называют запаздывающей (отстающей). Хотя отдельные фрагменты образуются в направлении 5″ → 3″, в целом эта цепь растет в направлении 3″ → 5″.
В виду того, что от локуса oriкак правило начинаются две репликационные вилки, идущие в противоположных направлениях, синтез лидирующих цепей в них идет на разных цепях материнской ДНК.
Конечным результатом процесса репликации является образование двух молекул ДНК, нуклеотидная последовательность которых идентична таковой в материнской двойной спирали ДНК.
Рассмотренная последовательность событий, происходящих в ходе репликативного синтеза, предполагает участие целой системы ферментов: хеликазы, топоизомеразы, дестабилизирующих белков, ДНК-полимеразы и других, совместно действующих в области репликационной вилки.
Репликация ДНК у про- и эукариот в основных чертах протекает сходно, однако, скорость синтеза у эукариот (около 100 нуклеотидов/с) на порядок ниже, чем у прокариот (1000 нуклеотидов/с). Причиной этого может быть образование ДНК эукариот достаточно прочных соединений с белками, что затрудняет ее деспирализацию, необходимую для осуществления репликативного синтеза.
Фрагмент ДНК от точки начала репликации до точки ее окончания образует единицу репликации — репликон. Однажды начавшись в точке начала, репликация продолжается до тех пор, пока весь репликон не будет дуплицирован. Эукариотические хромосомы содержат большое число репликонов. В связи с этим удвоение молекулы ДНК, расположенной вдоль эукариотической хромосомы, начинается в нескольких точках. В разных репликонах удвоение может идти в разное время или одновременно.
Уотсон и Крик показали, что образование водородных связей и регулярной двойной спирали возможно только тогда, когда более крупное пуриновое основание аденин (А) в одной цепи имеет своим партнером в другой цепи меньшее по размерам пиримидиновое основание тимин (Т), а гуанин (Г) связан с цитозином (Ц). Эту закономерность можно представить следующим образом:Соответствие А«Т и Г«Ц называют правилом комплементарности, а сами цепи- комплементарными. Согласно этому правилу, содержание аденина в ДНК всегда равно содержанию тимина, а количество гуанина количеству цитозина. Следует отметить, что две цепи ДНК, различаясь химически, несут одинаковую информацию, поскольку вследствие комплементарности одна цепь однозначно задает другую.
Структура РНК менее упорядочена. Обычно это одноцепочечная молекула, хотя РНК некоторых вирусов состоит из двух цепей. Но даже такая РНК более гибка, чем ДНК. Некоторые участки в молекуле РНК взаимно комплементарны и при изгибании цепи спариваются, образуя двухцепочечные структуры (шпильки). В первую очередь это относится к транспортным РНК (тРНК). Некоторые основания в тРНК подвергаются модификации уже после синтеза молекулы. Например, иногда происходит присоединение к ним метильных групп.
ФУНКЦИЯ НУКЛЕИНОВЫХ КИСЛОТ Одна из основных функций нуклеиновых кислот состоит в детерминации синтеза белков. Информация о структуре белков, закодированная в нуклеотидной последовательности ДНК, должна передаваться от одного поколения к другому, и поэтому необходимо ее безошибочное копирование, т.е. синтез точно такой же же молекулы ДНК (репликация).Репликация и транскрипция. С химической точки зрения синтез нуклеиновой кислоты это полимеризация, т.е. последовательное присоединение строительных блоков. Такими блоками служат нуклеозидтрифосфаты; реакцию можно представить следующим образом: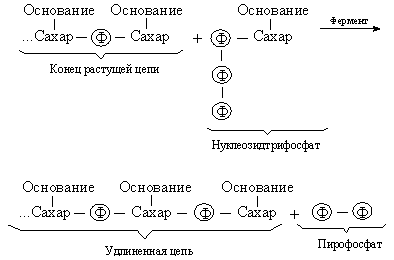 Энергия, необходимая для синтеза, высвобождается при отщеплении пирофосфата, а катализируют реакцию особые ферменты ДНК-полимеразы.
Энергия, необходимая для синтеза, высвобождается при отщеплении пирофосфата, а катализируют реакцию особые ферменты ДНК-полимеразы.
В результате такого синтетического процесса мы получили бы полимер со случайной последовательностью оснований. Однако большинство полимераз работает только в присутствии уже существующей нуклеиновой кислоты матрицы, диктующей, какой именно нуклеотид присоединится к концу цепи. Этот нуклеотид должен быть комплементарен соответствующему нуклеотиду матрицы, так что новая цепь оказывается комплементарной исходной. Используя затем комплементарную цепь в качестве матрицы, мы получим точную копию оригинала.
ДНК состоит из двух взаимно комплементарных цепей. В ходе репликации они расходятся, и каждая из них служит матрицей для синтеза новой цепи:
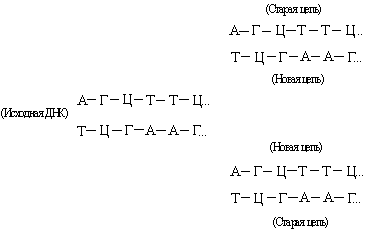 Так образуются две новые двойные спирали с той же последовательностью оснований, что и у исходной ДНК. Иногда в процессе репликации происходит «сбой», и возникают мутации (см. также НАСЛЕДСТВЕННОСТЬ).В результате транскрипции ДНК образуются клеточные РНК (мРНК, рРНК и тРНК):
Так образуются две новые двойные спирали с той же последовательностью оснований, что и у исходной ДНК. Иногда в процессе репликации происходит «сбой», и возникают мутации (см. также НАСЛЕДСТВЕННОСТЬ).В результате транскрипции ДНК образуются клеточные РНК (мРНК, рРНК и тРНК):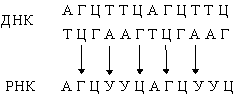 Они комплементарны одной из цепей ДНК и являются копией другой цепи, за исключением того, что место тимина у них занимает урацил. Таким способом можно получить множество РНК-копий одной из цепей ДНК.В нормальной клетке передача информации осуществляется только в направлении ДНК® ДНК и ДНК® РНК. Однако в клетках, инфицированных вирусом, возможны и другие процессы: РНК® РНК и РНК® ДНК. Генетический материал многих вирусов представлен молекулой РНК, обычно одноцепочечной. Проникнув в клетку-хозяина, эта РНК реплицируется с образованием комплементарной молекулы, на которой, в свою очередь, синтезируется множество копий исходной вирусной РНК:
Они комплементарны одной из цепей ДНК и являются копией другой цепи, за исключением того, что место тимина у них занимает урацил. Таким способом можно получить множество РНК-копий одной из цепей ДНК.В нормальной клетке передача информации осуществляется только в направлении ДНК® ДНК и ДНК® РНК. Однако в клетках, инфицированных вирусом, возможны и другие процессы: РНК® РНК и РНК® ДНК. Генетический материал многих вирусов представлен молекулой РНК, обычно одноцепочечной. Проникнув в клетку-хозяина, эта РНК реплицируется с образованием комплементарной молекулы, на которой, в свою очередь, синтезируется множество копий исходной вирусной РНК: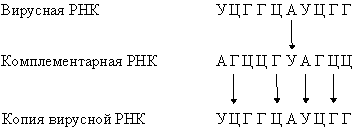 Вирусная РНК может транскрибироваться ферментом- обратной транскриптазой- в ДНК, которая иногда включается в хромосомную ДНК клетки-хозяина. Теперь эта ДНК несет вирусные гены, и после транскрипции в клетке может появиться вирусная РНК. Таким образом, спустя длительное время, в течение которого никакого вируса в клетке не обнаруживается, он снова в ней появится без повторного заражения. Вирусы, генетический материал которых включается в хромосому клетки-хозяина, часто являются причиной рака.
Вирусная РНК может транскрибироваться ферментом- обратной транскриптазой- в ДНК, которая иногда включается в хромосомную ДНК клетки-хозяина. Теперь эта ДНК несет вирусные гены, и после транскрипции в клетке может появиться вирусная РНК. Таким образом, спустя длительное время, в течение которого никакого вируса в клетке не обнаруживается, он снова в ней появится без повторного заражения. Вирусы, генетический материал которых включается в хромосому клетки-хозяина, часто являются причиной рака.
Молекулярнаягенетика–разделгенетики, который занимается изучениемнаследственности на молекулярномуровне.
Нуклеиновыекислоты. Репликация ДНК. Реакцииматричного синтеза
Нуклеиновыекислоты (ДНК, РНК) были открыты в 1868 годушвейцарским биохимиком И.Ф. Мишером.Нуклеиновые кислоты – линейныебиополимеры, состоящие из мономеров –нуклеотидов.
ДНК– структура и функции
Химическуюструктуру ДНК расшифровали в 1953 г.американский биохимик Дж. Уотсон ианглийский физик Ф. Крик.
Общаяструктура ДНК.МолекулаДНК состоит из 2 цепей, которые закрученыв спираль (рис. 11) одна вокруг другой ивокруг общей оси. Молекулы ДНК могутсодержать от 200 до 2х10 8пар нуклеотидов. Вдоль спирали молекулыДНК соседние нуклеотиды располагаютсяна расстоянии 0,34 нм друг от друга. Полныйоборот спирали включает 10 пар нуклеотидов.Его длина составляет 3,4 нм.
Рис.11.Схема строения ДНК (двойная спираль)
Полимерностьмолекулы ДНК.МолекулаДНК – биоплоимер состоит из сложныхсоединений – нуклеотидов.
Строениенуклеотида ДНК.НуклеотидДНК состоит из 3 звеньев: одно из азотистыхоснований (аденин, гуанин, цитозин,тимин); дезокисирибоза (моносахарид);остаток фосфорной кислоты (рис. 12).
Различают 2 группыазотистых оснований:
пуриновые – аденин (А), гуанин (Г), содержащие два бензольных кольца;
пиримидиновые – тимин (Т), цитозин (Ц), содержащие одно бензольное кольцо.
Всостав ДНК входят следующие видынуклеотидов: адениновый(А); гуаниновый (Г); цитозиновый (Ц);тиминовый (Т).Названиянуклеотидов соответствуют названиямазотистых оснований, входящих в ихсостав: адениновый нуклеотид азотистоеоснование аденин; гуаниновый нуклеотид азотистое основание гуанин; цитозиновыйнуклеотид азотистое основание цитозин;тиминовый нуклеотид азотистое основаниетимин.
Соединениедвух цепей ДНК в одну молекулу
НуклеотидыА, Г, Ц и Т одной цепи соединенысоответственно с нуклеотидами Т, Ц, Г иА другой цепи водороднымисвязями.Между А и Т формируется две водородныесвязи, а между Г и Ц – три водородныесвязи (А=Т, Г≡Ц).
Парыоснований (нуклеотидов) А – Т и Г – Цназывают комплементарными, т. е. взаимносоответствующими. Комплементарность– это химическое и морфологическоесоответствие нуклеотидов друг другу впарных цепочках ДНК.
5’ 3’
1 2 3
3’ 5’
Рис.12 Участокдвойной спирали ДНК. Строение нуклеотида(1– остаток фосфорной кислоты; 2–дезоксирибоза; 3– азотистое основание).Соединение нуклеотидов с помощьюводородных связей.
Цепив молекуле ДНК антипараллельны,т. е. направлены в противоположныестороны, так что 3’- конец одной цепирасполагается напротив 5’- конца другойцепи. Генетическая информация в ДНКзаписана в направлении от 5’ конца к 3’концу. Эта нить называется смысловойДНК,
посколькуздесь расположены гены. Вторая нить –3’–5’ служит эталоном хранениягенетической информации.
Cоотношениемежду числом разных оснований в ДНКустановлено Э. Чаргаффом в 1949 г. Чаргаффвыявил, что у ДНК различных видовколичество аденина равно количествутимина, а количество гуанина – количествуцитозина.
ПравилоЭ. Чаргаффа:
в молекуле ДНК количество A (адениновых) нуклеотидов всегда равно количеству Т (тиминовых) нуклеотидов или отношение ∑ А к ∑ Т=1. Сумма Г (гуаниновых) нуклеотидов равна сумме Ц (цитозиновых) нуклеотидов или отношение ∑ Г к ∑ Ц=1;
сумма пуриновых оснований (А+Г) равна сумме пиримидиновых оснований (Т+Ц) или отношение ∑ (А+Г) к ∑ (Т+Ц)=1;
Способсинтеза ДНК – репликация.Репликация – это процесс самоудвоениямолекулы ДНК, осуществляемый в ядре подконтролем ферментов. Самоудовоениемолекулы ДНК происходит наоснове комплементарности– строгого соответствия нуклеотидовдруг другу в парных цепочках ДНК. Вначале процесса репликации молекулаДНК раскручивается (деспирализуется)на определенном участке (рис. 13), приэтом освобождаются водородные связи.На каждой из цепей, образовавшихся послеразрыва водородных связей, при участиифермента ДНК-полимиразы,синтезируется дочерняя цепь ДНК.Материалом для синтеза служат свободныенуклеотиды, содержащиеся в цитоплазмеклеток. Эти нуклеотиды выстраиваютсякомплементарно нуклеотидам двухматеринских цепей ДНК. ФерментДНК-полимеразаприсоединяет комплементарные нуклеотидык матричной цепи ДНК. Например, кнуклеотиду Аматричной цепи полимераза присоединяетнуклеотид Ти, соответственно, к нуклеотиду Г –нуклеотид Ц (рис. 14). Сшивание комплементарныхнуклеотидов происходит с помощьюфермента ДНК-лигазы.Так путем самоудвоения синтезируютсядве дочерние цепи ДНК.
Образовавшиесядве молекулы ДНК из одной молекулы ДНКпредставляют собой полуконсервативнуюмодель,поскольку состоят из старой материнскойи новой дочерней цепей и являются точнойкопией материнской молекулы (рис. 14).Биологический смысл репликациизаключается в точной передаченаследственной информации от материнскоймолекулы к дочерней.

Рис.13.Деспирализация молекулы ДНК с помощьюфермента
1
Рис.14.Репликация – образование двух молекулДНК из одной молекулы ДНК: 1 – дочерняямолекула ДНК; 2 – материнская (родительская)молекула ДНК.
ФерментДНК-полимераза может двигаться вдольцепи ДНК только в направлении 3’ –>5’. Поскольку комплементарные цепи вмолекуле ДНК направлены в противоположныестороны, и фермент ДНК-полимераза можетдвигаться вдоль цепи ДНК только внаправлении 3’–>5’, то и синтез новыхцепей идет антипараллельно (попринципу антипараллельности).
Местолокализации ДНК.ДНК содержится в ядре клетки, в матриксемитохондрий и хлоропластов.
КоличествоДНК в клетке постоянно и составляет6,6х10 -12г.
Функции ДНК:
Хранение и передача в ряду поколений генетической информации молекулам и — РНК;
Структурная. ДНК является структурной основой хромосом (хромосома на 40% состоит из ДНК).
ВидоспецифичностьДНК.Нуклеотидный состав ДНК служит критериемвида.
РНК, строение ифункции.
Общаяструктура.
РНК– линейный биополимер, состоящий изодной полинуклеотидной цепи. Различаютпервичную и вторичную структуры РНК.Первичная структура РНК представляетсобой одноцепочечную молекулу, авторичная структура имеет форму крестаи характерна для т- РНК.
Полимерностьмолекулы РНК.Молекула РНК может включать от 70нуклеотидов до 30 000 нуклеотидов.Нуклеотиды, входящие в состав РНК,следующие: адениловый (А), гуаниловый(Г), цитидиловый (Ц), урациловый (У). Всоставе РНК тиминовый нуклеотид замещенна урациловый (У).
Строение нуклеотидаРНК.
Нуклеотид РНКвключает 3 звена:
азотистое основание (аденин, гуанин, цитозин, урацил);
моносахарид – рибоза (в рибозе присутствует кислород при каждом атоме углерода);
остаток фосфорной кислоты.
Способсинтеза РНК – транскрипция.Транскрипция, как и репликация, – реакцияматричного синтеза. Матрицей являетсямолекула ДНК. Реакция протекает попринципу комплементарности на однойиз цепей ДНК (рис. 15). Процесс транскрипцииначинается с деспирализации молекулыДНК на определенном участке. Натранскрибируемой цепи ДНК имеетсяпромотор –группануклеотидов ДНК, с которой начинаетсясинтез молекулы РНК. К промоторуприсоединяется фермент РНК-полимераза.Фермент активизирует процесс транскрипции.По принципу комплементарностидостраиваются нуклеотиды, поступающиеиз цитоплазмы клетки к транскрибируемойцепи ДНК. РНК-полимераза активизируетвыстраивание нуклеотидов в одну цепьи формирование молекулы РНК.
Впроцессе транскрипции выделяют четырестадии: 1) связывание РНК-полимеразы спромотором; 2) начало синтеза (инициация);3) элонгация – рост цепи РНК, т. е.происходит последовательное присоединениенуклеотидов друг к другу; 4) терминация– завершение синтеза и-РНК.

Рис.15.Схема транскрипции
1– молекула ДНК (двойная цепочка); 2 –молекула РНК; 3–кодоны; 4– промотор.
В1972 г. американские ученые – вирусологХ.М. Темин и молекулярный биолог Д.Балтимор на вирусах в опухолевых клеткахоткрыли обратную транскрипцию. Обратнаятранскрипция– переписывание генетической информациис РНК на ДНК. Процесс протекает с помощьюфермента обратнойтранскриптазы.
Виды РНК пофункции
Информационная, или матричная РНК (и-РНК, или м-РНК) переносит генетическую информацию с молекулы ДНК к месту синтеза белка – в рибосому. Синтезируется в ядре при участии фермента РНК-полимеразы. Она составляет 5% от всех видов РНК клетки. и- РНК включает от 300 нуклеотидов до 30 000 нуклеотидов (самая длинная цепь среди РНК).
Транспортная РНК (т-РНК) транспортирует аминокислоты к месту синтеза белка,– в рибосому. Имеет форму креста (рис. 16) и состоит из 70 – 85 нуклеотидов. Ее количество в клетке составляет 10-15 % РНК клетки.

Рис.16. Схемастроения т-РНК: А–Г – пары нуклеотидов,соединенные с помощью водородных связей;Д – место прикрепления аминокислоты(акцепторный участок); Е – антикодон.
3.Рибосомная РНК (р-РНК) синтезируется вядрышке и входит в состав рибосом.Включает примерно 3000 нуклеотидов.Составляет 85% РНК клетки. Этот вид РНКсодержатся в ядре, в рибосомах, наэндоплазматической сети, в хромосомах,в матриксе митохондрий, а также впластидах.
Основыцитологии. Решение типовых задач
Задача 1
Сколькотиминовых и адениновых нуклеотидовсодержится в ДНК, если в ней обнаружено50 цитозиновых нуклеотидов, что составляет10% от всех нуклеотидов.
Решение.По правилу комплементарности в двойнойцепи ДНК цитозин всегда комплемпентаренгуанину. 50 цитозиновых нуклеотидовсоставляют 10%, следовательно, согласноправилу Чаргаффа, 50 гуаниновых нуклеотидовтакже составляют 10%, или (если ∑Ц =10%, тои ∑Г =10%).
Суммапары нуклеотидов Ц + Г равна 20%
Суммапары нуклеотидов Т + А = 100% – 20 % (Ц + Г) =80 %
Длятого, чтобы узнать, сколько тиминовыхи адениновых нуклеотидов содержится вДНК, нужно составить следующую пропорцию:
50цитозиновых нуклеотидов → 10 %
Х (Т + А) →80 %
Х= 50х80:10=400 штук
Согласноправилу Чаргаффа ∑А= ∑Т, следовательно∑А=200 и ∑Т=200.
Ответ:количествотиминовых, как и адениновых нуклеотидовв ДНК, равно 200.
Задача 2
Тиминовыенуклеотиды в ДНК составляют 18% от общегоколичества нуклеотидов. Определитепроцент остальных видов нуклеотидов,содержащихся в ДНК.
Решение.∑Т=18%. Согласно правилу Чаргаффа ∑Т=∑А,следовательно на долю адениновыхнуклеотидов также приходится 18 % (∑А=18%).
Суммапары нуклеотидов Т+А равна 36 % (18 % + 18 % =36 %). На пару нуклеотидов Ги Ц приходится:Г+Ц=100 % –36 %=64 %. Поскольку гуанин всегдакомплементарен цитозину, то их содержаниев ДНК будет равным,
т.е. ∑ Г= ∑Ц=32%.
Ответ:содержание гуанина, как и цитозина,составляет 32 %.
Задача 3
20цитозиновых нуклеотидов ДНК составляют10% от общего количества нуклеотидов.Сколько адениновых нуклеотидов содержитсяв молекуле ДНК?
Решение.В двойной цепочке ДНК количество цитозинаравно количеству гуанина, следовательно,их сумма составляет: Ц+Г=40 нуклеотидов.Находим общее количество нуклеотидов:
20 цитозиновых нуклеотидов → 10 %
Х(общее количество нуклеотидов) →100 %
Х=20х100:10=200штук
А+Т=200 – 40=160 штук
Так как аденинкомплементарен тимину, то их содержаниебудет равным,
т.е. 160 штук: 2=80 штук, или ∑А=∑Т=80.
Ответ:в молекуле ДНК содержится 80 адениновыхнуклеотидов.
Задача 4
Допишитенуклеотиды правой цепи ДНК, если известнынуклеотиды ее левой цепи: АГА – ТАТ –ГТГ – ТЦТ
Решение.Построение правой цепи ДНК по заданнойлевой цепи производится по принципукомплементарности – строгого соответствиянуклеотидов друг другу: аденонивый –тиминовый (А–Т), гуаниновый – цитозиновый(Г–Ц). Поэтому нуклеотиды правой цепиДНК должны быть следующие: ТЦТ – АТА –ЦАЦ – АГА.
Ответ:нуклеотиды правой цепи ДНК: ТЦТ – АТА– ЦАЦ – АГА.
Задача 5
Запишитетранскрипцию, если транскрибируемаяцепочка ДНК имеет следующий порядокнуклеотидов: АГА – ТАТ – ТГТ – ТЦТ.
Решение.Молекула и-РНК синтезируется по принципукомплеиентарности на одной из цепеймолекулы ДНК. Нам известен порядокнуклеотидов в транскрибируемой цепиДНК. Следовательно, надо построитькомплементарную цепь и-РНК. Следуетпомнить, что вместо тимина в молекулуРНК входит урацил. Следовательно:
ЦепьДНК: АГА – ТАТ – ТГТ – ТЦТ
Цепьи-РНК: УЦУ – АУА –АЦА –АГА.
Ответ:последовательность нуклеотидов и-РНКследующая: УЦУ – АУА – АЦА –АГА.
Задача 6
Запишитеобратную транскрипцию, т. е. постройтефрагмент двухцепочечной молекулы ДНКпо предложенному фрагменту и-РНК, еслицепочка и- РНК имеет следующуюпоследовательность нуклеотидов:
ГЦГ– АЦА – УУУ – УЦГ – ЦГУ – АГУ – АГА
Решение.Обратная транскрипция – это синтезмолекулы ДНК на основе генетическогокода и-РНК. Кодирующая молекулу ДНКи-РНК имеет следующий порядок нуклеотидов:ГЦГ – АЦА – УУУ – УЦГ – ЦГУ – АГУ –АГА. Комплементарная ей цепочка ДНК:ЦГЦ – ТГТ – ААА – АГЦ – ГЦА – ТЦА –ТЦТ. Вторая цепочка ДНК:ГЦГ–АЦА–ТТТ–ТЦГ–ЦГТ–АГТ–АГА.
Ответ:в результате обратной транскрипциисинтезированы две цепочки молекулыДНК: ЦГЦ – ТГТ – ААА – АГЦ – ГЦА – ТЦАи ГЦГ–АЦА–ТТТ–ТЦГ–ЦГТ–АГТ–АГА.
Генетическийкод. Биосинтез белка.
Ген– участок молекулы ДНК, содержащийгенетическую информацию о первичнойструктуре одного определенного белка.
Экзон-интроннаяструктура генаэукариот
промотор – участок ДНК (длиной до 100 нуклеотидов), к которому присоединяется фермент РНК-полимераза, необходимый для осуществления транскрипции;
2)регуляторнаязона – зона,влияющая на активность гена;
3)структурнаячасть гена– генетическая информация о первичнойструктуре белка.
Последовательностьнуклеотидов ДНК, несущая генетическуюинформацию о первичной структуре белка– экзон.Они также входят в состав и-РНК.Последовательность нуклеотидов ДНК,не несущая генетическую информацию опервичной структуре белка –интрон. Онине входят в состав и-РНК. В ходе транскрипциис помощью специальных ферментовпроисходит вырезание копий интроновиз и-РНК и сшивание копий экзонов приобразовании молекулы и-РНК (рис. 20). Этотпроцесс называется сплайсинг.

Рис.20.Схема сплайсинга (формирование зрелойи-РНК у эукариот)
Генетическийкод –системапоследовательности нуклеотидов вмолекуле ДНК, или и-РНК, котораясоответствует последовательностиаминокислот в полипептидной цепи.
Свойствагенетического кода:
Триплетность(АЦА – ГТГ – ГЦГ…)
Генетическийкод являетсятриплетным,таккак каждая из 20 аминокислот кодируетсяпоследовательностью трех нуклеотидов(триплетом,кодоном).
Существует64 вида триплетов нуклеотидов (4 3 =64).
Однозначность (специфичность)
Генетическийкод является однозначным, так каккаждыйотдельный триплет нуклеотидов (кодон)кодирует только одну аминокислоту, илиодин кодон всегда соответствует однойаминокислоте (таблица 3).
Множественность (избыточность, или вырожденность)
Однаи та же аминокислота может кодироватьсянесколькими триплетами (от 2 до 6), т. к.белокобразующих аминокислот –20, атриплетов – 64.
Непрерывность
Считываниегенетической информации происходит водном направлении, слева направо. Еслипроизойдет выпадение одного нуклеотида,то при считывании его место займетближайший нуклеотид из соседнеготриплета, что приведет к изменениюгенетической информации.
Универсальность
Генетическийкод характерен для всех живых организмов,и одинаковые триплеты кодируют одну иту же аминокислоту у всех живых организмов.
Имеет стартовые и терминальные триплеты (стартовый триплет – АУГ, терминальные триплеты УАА, УГА, УАГ). Эти виды триплетов не кодируют аминокислоты.
Неперекрываемость (дискретность)
Генетическийкод является неперекрывающимся, таккак один и тот же нуклеотид не можетвходить одновременно в состав двухсоседних триплетов. Нуклеотиды могутпринадлежать только одному триплету,а если переставить их в другой триплет,то произойдет изменение генетическойинформации.
Таблица3 – Таблица генетического кода
Основания кодонов
Примечание:сокращенные названия аминокислот даныв соответствии с международнойтерминологией.
Биосинтезбелка
Биосинтезбелка – видпластического обменавеществ в клетке, происходящий в живыхорганизмах под действием ферментов.Биосинтезу белка предшествуют реакцииматричного синтеза (репликация – синтезДНК; транскрипция – синтез РНК; трансляция– сборка молекул белка на рибосомах).В процессе биосинтеза белка выделяют2 этапа:
транскрипция
трансляция
Входе транскрипции генетическаяинформация, заключенная в ДНК, находящейсяв хромосомах ядра, передается молекулеРНК. По завершении процесса транскрипциии-РНК выходит в цитоплазму клетки черезпоры в мембране ядра, располагаетсямежду 2 субъединицами рибосомы и участвуетв биосинтезе белка.
Трансляция– процесс перевода генетического кодав последовательность аминокислот.Трансляцияосуществляется в цитоплазме клетки нарибосомах, которые располагаются наповерхности ЭПС (эндоплазматическойсети). Рибосомы – сферические гранулы,диаметром, в среднем, 20 нм, состоящие избольшой и малой субъединиц. Молекулаи-РНК располагается между двумясубъединицами рибосомы. В процессетрансляции участвуют аминокислоты,АТФ, и-РНК, т-РНК, фермент амино-ацилт-РНК-синтетаза.
Кодон– участок молекулы ДНК, или и-РНК,состоящий из трех последовательнорасположенных нуклеотидов, кодирующийодну аминокислоту.
Антикодон– участокмолекулы т-РНК, состоящий из трехпоследовательно расположенных нуклеотидови комплементарный кодону молекулыи-РНК. Кодоны комплементарны соответствующимантикодонам и соединяются с ними спомощью водородных связей (рис. 21).
Синтезбелка начинается со стартовогокодона АУГ.От него рибосома
перемещаетсяпо молекуле и-РНК, триплет за триплетом.Аминокислоты поступают по генетическомукоду. Встраивание их в полипептиднуюцепь на рибосоме происходит с помощьют-РНК. Первичная структура т-РНК (цепочка)переходит во вторичную структуру,напоминающую по форме крест, и при этомв ней сохраняется комплементарностьнуклеотидов. В нижней части т-РНК имеетсяакцепторный участок, к которомуприсоединяется аминокислота (рис.16).Активизация аминокислоты осуществляетсяпри помощи фермента аминоацилт-РНК-синтетазы.Суть этого процесса состоит в том, чтоданный фермент взаимодействует саминокислотой и с АТФ. При этом формируетсятройной комплекс, представленный даннымферментом, аминокислотой и АТФ.Аминокислота обогащается энергией,активизируется, приобретает способностьобразовывать пептидные связи с соседнейаминокислотой. Без процесса активизацииаминокислоты полипептидная цепь изаминокислт сформироваться не может.
Впротивоположной, верхней части молекулыт-РНК содержится триплет нуклеотидовантикодон,с помощью которого т-РНК прикрепляетсяк комплементарному ему кодону (рис. 22).
Перваямолекула т-РНК, с присоединенной к нейактивизированной аминокислотой, своимантикодоном прикрепляется к кодонуи-РНК, и в рибосоме оказывается однааминокислота. Затем прикрепляетсявторая т-РНК своим антикодоном ксоответствующему кодону и-РНК. При этомв рибосоме оказываются уже 2 аминокислоты,между которыми формируется пептиднаясвязь. Первая т-РНК покидает рибосому,как только отдаст аминокислоту вполипептидную цепь на рибосоме. Затемк дипептиду присоединяется 3-я аминокислота,ее приносит третья т-РНК и т. д. Синтезбелка останавливается на одном изтерминальных кодонов – УАА, УАГ, УГА(рис. 23).

1– кодони-РНК;кодоныUCG –УЦГ;CUA –ЦУА;CGU –ЦГУ;
2–антикодон т-РНК; антикодон GAT – ГАТ
Рис.21.Фаза трансляции: кодон и-РНК притягиваетсяк антикодону т-РНК соответствующимикомплементарными нуклеотидами(основаниями)
