1. В митохондриях происходит синтез ATP (см. Окислительное фосфорилирование)
PH межмембранного пространства ~4, pH матрикса ~8 | содержание белков в м: 67% - матрикс, 21% -наруж м-на, 6% - внутр м-на и 6% - в межм-ном пр-ве
Хандриома
– единая система митохондрий
наружная м-на: порины-поры позволяют проходить до 5 kD | внутренняя м-на: кардиолипин-делает непроницаемой м-ну для ионов |
межм-ное пр-во: группы ферментов фосфорилируют нуклеотиды и сахара нуклеотидов
внутренняя м-на:
матрикс: метаболические ферменты – окисление липидов, окисление углеводов, цикла трикарбоновых к-т, цикла Кребса
Происхождение от бактерий: амеба Pelomyxa palustris единств из эукариот не содержит м., живет в симбиозе с аэробными бактериями | собственная ДНК | схожие с бактериями оx процессы
16569пн=13белков,22тРНК,2pРНК | гладкая внешняя м-на (порины – проницаемость белков до 10 кДа) складчатая внутренняя (кристы) м-на (75% -белков: транспортные белки-переносчики, ф-ты, компаненты дыхат. цепи и АТФ-синтаза, кардиолипин) матрикс (обогащен ф-тами цитратного цикла) межм-ное пр-во
Митохондрии - органеллы энергообеспечения
метаболических процесов в клетке.
Размеры их варьируют от 0,5 до 5-7 мкм,
количество в клетке составляет от 50 до
1000 и более. В гиалоплазме митохондрии
распределены обычно диффузно, однако
в специализированных клетках сосредоточены
в тех участках, где имеется наибольшая
потребность в энергии. Например, в
мышечных клетках и симпластах большие
количества митохондрий сосредоточены
вдоль рабочих элементов - сократительных
фибрилл. В клетках, функции которых
сопряжены с особо высокими энергозатратами,
митохондрии образуют множественные
контакты, объединяясь в сеть, или
кластеры (кардиомиоциты и симпласты
скелетной мышечной ткани). В клетке
митохондрии выполняют функцию дыхания.
Клеточное дыхание - это последовательность
реакций, с помощью которых клетка
использует энергию связей органических
молекул для синтеза макроэргических
соединений типа АТФ. Образующиеся
внутри митохондрии молекулы АТФ
переносятся наружу, обмениваясь на
молекулы АДФ, находящиеся вне митохондрии.
В живой клетке митохондрии могут
передвигаться с помощью элементов
цитоскелета. На ультрамикроскопическом
уровне стенка митохондрии состоит из
двух мембран - наружной и внутренней.
Наружная мембрана имеет относительно
ровную поверхность, внутренняя -
образует направленные в центр складки,
или кристы. Между наружной и внутренней
мембранами возникает неширокое (около
15 нм) пространство, которое называется
наружной камерой митохондрии; внутренняя
мембрана ограничивает внутреннюю
камеру. Содержимое наружной и внутренней
камер митохондрии различно, и так же,
как и сами мембраны, существенно
отличается не только по рельефу
поверхности, но и по ряду биохимических
и функциональных признаков. Наружная
мембрана по химическому составу и
свойствам близка к другим внутриклеточным
мембранам и плазмолемме.
Ее характеризует
высокая проницаемость, благодаря
наличию гидрофильных белковых каналов.
Эта мембрана имеет в своем составе
рецепторные комплексы, распознающие
и связывающие вещества, поступающие в
митохондрию. Ферментный спектр наружной
мембраны небогат: это ферменты метаболизма
жирных кислот, фосфолипидов, липидов
и др. Главной функцией наружной
мембраны митохондрии является
отграничение органеллы от гиалоплазмы
и транспорт необходимых для осуществления
клеточного дыхания субстратов. Внутренняя
мембрана митохондрий в большинстве
клеток тканей различных органов
формирует кристы в виде пластин
(ламеллярные кристы), что значительно
увеличивает площадь поверхности
внутренней мембраны. В последней 20-25 %
всех белковых молекул составляют
ферменты дыхательной цепи и окислительного
фосфорилирования. В эндокринных клетках
надпочечников и половых желез митохондрии
участвуют в синтезе стероидных гормонов.
В этих клетках митохондрии имеют кристы
в виде трубочек (тубул), упорядоченно
расположенных в определенном направлении.
Поэтому кристы митохондрий в
стероидпродуцирующих клетках названных
органов именуются тубулярными. Матрикс
митохондрии, или содержимое внутренней
камеры, представляет собой гелеобразную
структуру, содержащую около 50 % белков.
Осмиофильные тельца, описанные при
электронной микроскопии, - это резервы
кальция. Матрикс содержит ферменты
цикла лимонной кислоты, катализирующие
окисление жирных кислот, синтез рибосом,
ферменты, участвующие в синтезе РНК и
ДНК. Общее число ферментов превышает
40. Помимо ферментов, матрикс митохондрии
содержит митохондриальную ДНК (митДНК)
и митохондриальные рибосомы. Молекула
митДНК имеет кольцевидную форму.
Возможности внутримитохондриального
белкового синтеза ограничены - здесь
синтезируются транспортные белки
митохондриальных мембран и некоторые
ферментные белки, участвующие в
фосфорилировании АДФ. Все остальные
белки митохондрии кодируются ядерной
ДНК, и их синтез осуществляется в
гиалоплазме, и в дальнейшем они
транспортируются в митохондрию.
Жизненный цикл митохондрий в клетке
короткий, поэтому природа наделила их
двойственной системой воспроизводства
- помимо деления материнской митохондрии,
возможно образование нескольких
дочерних органелл путем почкования.
Митохондрия.
Митохондрия
- состоящая из двух мембран органелла толщиной около 0,5 мкм.
Энергетическая станция клетки; основная функция - окисление органических соединений и использование, освобождающейся при их распаде энергии в синтезе молекул атф (универсальный источник энергии для всех биохимических процессов).
По своему строению они представляют собой цилиндрические органеллы, встречающиеся в эукариотической клетке в количестве от нескольких сот до 1-2 тысяч и занимающие 10-20 % её внутреннего объёма. Сильно варьируют так же размеры (от 1 до 70 мкм) и форма митохондрий. При этом ширина этих частей клетки относительно постоянна (0,5-1 мкм). Способны изменять форму. в зависимости от того, в каких участках клетки в каждый конкретный момент происходит повышенное потребление энергии, митохондрии способны перемещаться по цитоплазме в зоны наибольшего энергопотребления, используя для движения структуры клеточного каркаса эукариотической клетки.
Красавица митохондрия в 3д представлении)
Альтернативой множеству разрозненных небольших митохондрий, функционирующих независимо друг от друга и снабжающих атф небольшие участки цитоплазмы, является существование длинных и разветвлённых митохондрий, каждая из которых может энергетически обеспечивать отдалённые друг от друга участки клетки. вариантом такой протяжённой системы может также являться упорядоченное пространственное объединение множества митохондрий (хондриом или митохондрион), обеспечивающее их кооперативную работу.
Особенно сложно этот тип хондриома устроен в мышцах, где группы гигантских разветвлённых митохондрий связаны друг с другом с помощью межмитохондриальных контактов (ммк). Последние образованы плотно прилегающими друг к другу наружными митохондриальными мембранами, в результате чего межмембранное пространство в этой зоне имеет повышенную электронную плотность (много отрицательно заряженных частиц). Особенно обильно ммк представлены в клетках сердечных мышц, где они связывают множественные отдельные митохондрии в согласованную работающую кооперативную систему.
Структура.
Наружная мембрана.
Наружная мембрана митохондрии имеет толщину около 7 нм, не образует впячиваний и складок, и замкнута сама на себя. на наружную мембрану приходится около 7 % от площади поверхности всех мембран клеточных органелл. Основная функция - отграничение митохондрии от цитоплазмы. Наружная мембрана митохондрии состоит из двойного жирового слоя (как и у клеточной мембраны) и пронизывающих его белков. Белки и жиры в равных пропорциях по массе.
Особую роль играет порин
-
каналообразующий белок.
Он формирует в наружной мембране отверстия диаметром 2-3 нм, через которые могут проникать небольшие молекулы и ионы. Крупные молекулы могут пересекать наружную мембрану только посредством активного транспорта через транспортные белки митохондриальных мембран. Наружная мембрана митохондрии может взаимодействовать с мембраной эндоплазматического ретикулума; это играет важную роль в транспортировке липидов и ионов кальция.
Внутренняя мембрана.
Внутренняя мембрана образует многочисленные гребневидные складки -
кристы,
существенно увеличивающие площадь ее поверхности и, например, в клетках печени составляет около трети всех клеточных мембран. характерной чертой состава внутренней мембраны митохондрий является присутствие в ней кардиолопина
-
особый сложный жир, содержащего сразу четыре жирные кислоты и делающего мембрану абсолютно непроницаемой для протонов (положительно заряженных частиц).
Ещё одна особенность внутренней мембраны митохондрий - очень высокое содержание белков (до 70 % по весу), представленных транспортными белками, ферментами дыхательной цепи, а также крупными ферментами комплексами производящими атф. Внутренняя мембрана митохондрии в отличие от внешней не имеет специальных отверстий для транспорта мелких молекул и ионов; на ней, на стороне, обращенной к матриксу, располагаются особые молекулы ферменты производящие атф, состоящие из головки, ножки и основания. При прохождении через них протонов происходит создание атф.
В основании частиц, заполняя собой всю толщу мембраны, располагаются компоненты дыхательной цепи. наружная и внутренняя мембраны в некоторых местах соприкасаются, там находится специальный белок-рецептор, способствующий транспорту митохондриальных белков, закодированных в ядре, в матрикс митохондрии.
Матрикс.
Матрикс
- ограниченное внутренней мембраной пространство. В матриксе (розовом веществе) митохондрии находятся ферментные системы окисления пирувата жирных кислот, а так же ферменты типа трикарбоновых кислот (цикл дыхания клетки). Кроме того, здесь же находится митохондриальная днк, рнк и собственный белоксинтезирующий аппарат митохондрии.
пируваты (соли пировиноградной кислоты)
- важные химические соединения в биохимии. Они является конечным продуктом обмена веществ глюкозы в процессе ее расщепления.
Митохондриальная днк.
Несколько отличий от днк ядерной:
- митохондриальная днк – кольцевая, в отличии от ядерной днк, которая упакована в хромосомы.
- между различными эволюционными вариантами митохондриальной днк одного вида невозможен обмен сходными участками.
И поэтому вся молекула изменяется только путем медленного мутирования в течение тысячелетий.
- мутации кода в митохондриальных днк могут возникать независимо от ядерной днк.
Мутация ядерного кода днк возникает в основном при делении клетки, но митохондрии делятся независимо от клетки, и могут получать мутацию кода отдельно от ядерной днк.
- сама структура митохондриальной днк упрощена, т.к. многие составные процессы чтения днк утеряны.
- транспортные рнк имеют одинаковое строение. но рнк- митохондрий учавствуют только в синтезе митохондриальных белков.
Имея собственный генетический аппарат, митохондрия обладает и собственной белоксинтезирующей системой, особенностью которой в клетках животных и грибов являются очень маленькие рибосомы.
Функции.
Энергообразование.
Основной функцией митохондрий является синтез атф - универсальной формы химической энергии в любой живой клетке.
Данная молекула может образовываться двумя путями:
- путем реакции, в которых энергия освобождающаяся на определенных окислительных этапах брожения запасается в виде атф.
- благодаря энергии, выделяющейся при окислении органических веществв в процессе клеточного дыхания.
Митохондрии реализуют оба эти пути, первый из которых характерен для начальных процессов окисления и происходит в матриксе, а второй завершает процессы энергообразования и связан с кристами митохондрий.
При этом своеобразие митохондрий как энергообразующих органелл эукариотической клетки определяет именно второй путь генерации атф, получивший название «хемиосмотического сопряжения».
В целом весь процесс энергообразования в митохондриях может быть разбит на четыре основные стадии, первые две из которых протекают в матриксе, а две последние - на кристах митохондрий:
1) Превращение поступивших из цитоплазмы в митохондрию пируват (конечный продукт расщепления глюкозы) и жирных кислот в ацетил-коа;
ацетил-коа
– важное соединение в обмене веществ, используемое во многих биохимических реакциях. его главная функция – доставлять атомы углерода (с) с ацетил-группой (ch3 co) в цикл клеточного дыхания, чтобы те были окислены с выделением энергии.
клеточное дыхание
-
совокупность биохимических реакций, протекающих в клетках живых организмов, в ходе которых происходит окисление углеводов, жиров и аминокислот до углекислого газа и воды.
2) Окисление ацетил-соа в цикле клеточного дыхания, ведущее к образованию надн;
НАДН
–
кофермент, выполняет функцию переносчика электронов и водорода, которые принимает от окисляемых веществ.
3) Перенос электронов с надн на кислород по дыхательной цепи;
4) Образование атф в результате деятельности мембранного атф-создающего комплекса.
АТФ- синтетаза.
АТФ-синтетаза
–
станция по производству молекул АТФ.
В структурно-функциональном плане АТФ-синтетаза состоит из двух крупных фрагментов, обозначаемых символами F1 и F0. Первый из них (фактор сопряжения F1) обращён в сторону матрикса митохондрии и заметно выступает из мембраны в виде сферического образования высотой 8 нм и шириной 10 нм. Он состоит из девяти субъединиц, представленных пятью типами белков. Полипептидные цепи трёх субъединиц α и стольких же субъединиц β уложены в похожие по строению белковые глобулы, которые вместе образуют гексамер (αβ)3, имеющий вид слегка приплюснутого шара.
Субъединица
– это структурный и функциональный компонент какой либо частицы
Полипептиды
- органические соединения, содержащие от 6 до 80-90 аминокислотных остатков.
Глобула
– состояние макромолекул, в котором колебание звеньев мало.
Гексамер
– соединение содержащее 6 субъедениц.
Подобно плотно уложенным долькам апельсина, последовательно расположенные субъединицы α и β образуют структуру, характеризующуюся симметричность относительно угла поворота 120°. В центре этого гексамера находится субъединица γ, которая образована двумя протяжёнными полипептидными цепями и напоминает слегка деформированный изогнутый стержень длиной около 9 нм. При этом нижняя часть субъединицы γ выступает из шара на 3 нм в сторону мембранного комплекса F0. Также внутри гексамера находится минорная субъединица ε, связанная с γ. Последняя (девятая) субъединица обозначается символом δ и расположена на внешней стороне F1.
Минорная
– одиночная субъеденица.
Мембранная часть АТФ-синтетазы, представляет собой водо-отталкивающий белковый комплекс, пронизывающий мембрану насквозь и имеющий внутри себя два полуканала для прохождения протонов водорода. Всего в состав комплекса F0 входит одна белковая субъединица типа а
, две копии субъединицы b
, а также от 9 до 12 копий мелкой субъединицы c
. Субъединица а
(молекулярная масса 20 кДа) полностью погружена в мембрану, где образует шесть пересекающих её α-спиральных участков. Субъединица b
(молекулярная масса 30 кДа) содержит лишь один сравнительно короткий погружённый в мембрану α-спиральный участок, а остальная её часть заметно выступает из мембраны в сторону F1 и закрепляется за расположенную на её поверхности субъединицу δ. Каждая из 9-12 копий субъединицы c
(молекулярная масса 6-11 кДа) представляет собой сравнительно небольшой белок из двух водо-отталкивающих α-спиралей, соединённых друг с другом короткой водо-притягивающей петлёй, ориентированной в сторону F1, а все вместе образуют единый ансамбль, имеющий форму погружённого в мембрану цилиндра. Выступающая из комплекса F1 в сторону F0 субъединица γ как раз и погружена внутрь этого цилиндра и достаточно прочно зацеплена за него.
Таким образом, в молекуле АТФазы можно выделить две группы белковых субъединиц, которые могут быть уподоблены двум деталям мотора: ротору и статору.
«Статор»
неподвижен относительно мембраны и включает в себя шарообразный гексамер (αβ)3, находящуюся на его поверхности и субъединицу δ, а также субъединицы a
и b
мембранного комплекса F0.
Подвижный относительно этой конструкции «ротор»
состоит из субъединиц γ и ε, которые, заметно выступая из комплекса (αβ)3, соединяются с погружённым в мембрану кольцом из субъединиц c
.
Способность синтезировать АТФ - свойство единого комплекса F0F1, объедененного с переносом протонов водорода через F0 к F1, в последнем из которых как раз и расположены центры реакции, осуществляющие преобразование АДФ и фосфата в молекулу АТФ. Движущей же силой для работы АТФ-синтетазы является протонный (положительно заряженный) потенциал, создаваемый на внутренней мембране митохондрий в результате работы цепи электронного (отрицательно заряженного) транспорта.
Сила, приводящая в движение «ротор» АТФ-синтетазы, возникает при достижении разности потенциалов между наружной и внутренней сторонами мембраны > 220 10−3 Вольт и обеспечивается потоком протонов, протекающих через специальный канал в F0, расположенный на границе между субъединицами a
и c
. При этом путь переноса протонов включает в себя следующие структурные элементы:
1) Два расположенных на разных осях «полуканала», первый из которых обеспечивает поступление протонов из межмембранного пространства к существенно важным функциональным группам F0, а другой обеспечивает их выход в матрикс митохондрии;
2) Кольцо из субъединиц c
, каждая из которых в своей центральной части содержит протонируемую карбоксильную группу (COOH), способную присоединять H+ из межмембранного пространства и отдавать их через соответствующие протонные каналы. В результате периодических смещений субъединиц с
, обусловленных потоком протонов через протонный канал происходит поворот субъединицы γ, погружённой в кольцо из субъединиц с
.
Таким образом, объединяющая активность АТФ-синтетазы непосредственно связана с вращением её «ротора», при котором поворот субъединицы γ вызывает одновременное изменение конформации всех трёх объединяющих субъединиц β, что в конечном счёте и обеспечивает работу фермента. При этом в случае образования АТФ «ротор» крутится по часовой стрелке со скоростью четыре оборота в секунду, а само подобное вращение происходит точными скачками по 120°, каждый из которых сопровождается образованием одной молекулы АТФ.
Работа АТФ-синтетазы связана с механическими движениями её отдельных частей, что позволило отнести этот процесс к особому типу явлений, названных «вращательным катализом». Подобно тому, как электрический ток в обмотке электродвигателя приводит в движение ротор относительно статора, направленный перенос протонов через АТФ-синтетазу вызывает вращение отдельных субъединиц фактора сопряжения F1 относительно других субъединиц ферментного комплекса, в результате чего это уникальное энергообразующее устройство совершает химическую работу - синтезирует молекулы АТФ. В дальнейшем АТФ поступает в цитоплазму клетки, где расходуется на самые разнообразные энергозависимые процессы. Подобный перенос осуществляется специальным встроенным в мембрану митохондрий ферментом АТФ/АДФ-транслоказой.
АДФ-транслоказа
– пронизывающий внутреннюю мембрану белок, который обменивает вновь синтезированную АТФ на цитоплазматическую АДФ, что гарантирует сохранность фонда внутри митохондрий.
Митохондрии и наследственность.
ДНК митохондрий наследуются почти исключительно по материнской линии. Каждая митохондрия имеет несколько участков нуклеотидов в ДНК, идентичных во всех митохондриях (то есть в клетке много копий митохондриальных ДНК), что очень важно для митохондрий, неспособных восстанавливать ДНК от повреждений (наблюдается высокая частота мутаций). Мутации в митохондриальной ДНК являются причиной целого ряда наследственных заболеваний человека.
3д модель
Дисковери
С англ озвучкой
Немного о дыхании клетки и митохондрии на зарубежном языке
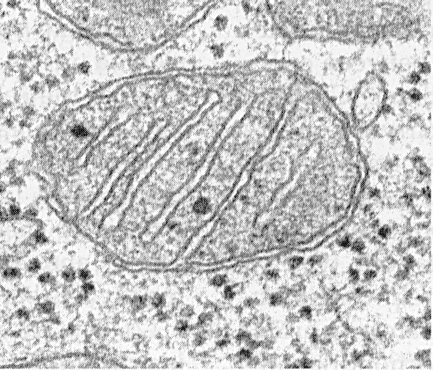
Структура строения
Каждая митохондрия состоит из наружной
и внутренней мембран
, между которыми находится межмембранное пространство (Рис
.7). Внутренняя мембрана образует складки - кристы
, обращенные внутрь митохондрии. Пространство, ограниченное внутренней мембраной, заполнено митохондриальным матриксом
, - мелкозернистым материалом различной электронной плотности.
Рис.7.
Наружная мембрана
митохондрий содержит много молекул специализированных транспортных белков (например, порин), что обеспечивает её высокую проницаемость, а также белки-рецепторы, распознающие белки, которые переносятся через обе мембраны митохондрий в особых точках их контакта – зонах слипания.
Внутренняя мембрана
митохондрий образует складки – кристы
, благодаря чему значительно увеличивается внутренняя поверхность митохондрий. В состав внутренней мембраны входят транспортные белки; ферменты дыхательной цепи и сукцинатдегидрогеназа; комплекс АТФ-синтетазы. На кристах имеются элементарные частицы (оксисомы
, или F1-частицы), состоящие из округлой головки (9 нм) и цилиндрической ножки. Именно на них происходит сопряжение процессов окисления и фосфорилирования (АДФ → АТФ).
Чаще всего кристы располагаются перпендикулярно длинной оси митохондрий и имеют пластинчатую
(ламеллярную
) форму. В клетках,синтезирующих стероидные гормоны, кристы имеют вид трубочек или пузырьков - тубулярно-везикулярные кристы
. В этих клетках ферменты стероидного синтеза частично локализуются на внутренней мембране митохондрий
Число и площадь крист отражает функциональную активность клеток: наибольшая площадь крист характерна, например, для митохондрий клеток сердечной мышцы, где потребность в энергии постоянно очень велика.
Митохондриальный матрикс –
мелкозернистое вещество, заполняющее полость митохондрии. Матрикс содержит несколько сотен ферментов: ферменты цикла Кребса, окисления жирных кислот, белкового
синтеза
. Здесь иногда встречаются митохондриальные гранулы
, а также локализуются митохондриальные ДНК, иРНК, тРНК, рРНК
и митохондриальные рибосомы.
Митохондриальные гранулы – частицы высокой электронной плотности диаметром 20-50 нм, содержащие ионы Са и Мg.
Митохондриальная ДНК имеет кольцевую форму и включает 37 генов. Генетическая информация митохондриальной ДНК обеспечивает синтез около 5-6% белков митохондрий (ферменты электрон-транспортной системы). Синтез других митохондриальных белков контролируется ДНК ядра. Наследование митохондриальной ДНК происходит только по материнской линии.
Повреждения митохондриальной ДНК в результате мутаций могут привести к развитию ряда патологий - митохондриальных цитопатий (синдромы Барта, Патерсона, МERRF (красных разорванных волокон) и др.).
ЛИЗОСОМЫ
– мембранные органеллы, которые обеспечивают внутриклеточное переваривание
(расщепление) макромолекул внеклеточного и внутриклеточного происхождения, и обновление компонентов клетки.
Морфологически лизосомы представляют собой округлые пузырьки, ограниченные мембраной и содержащие большое количество различных гидролаз (более 60 ферментов). Наиболее характерными ферментами лизосом являются: кислая фосфатаза (маркёр
лизосом), протеазы, нуклеазы, сульфатазы, липазы, гликозидазы. Все литические ферменты лизосом представляют собой кислые гидролазы
, т.е. оптимум их активности проявляется при рН≈5.
Мембрана лизосом (около 6 нм толщиной) обладает протонным насосом
, вызывающим закисление среды внутри органелл, обеспечивает диффузию низкомолекулярных продуктов переваривания макромолекул в гиалоплазму и препятствует утечке литических ферментов в гиалоплазму.
Повреждение мембраны приводит к разрушению клетки вследствие самопереваривания.
Лизосомы присутствуют во всех клетках. Особенно много лизосом в тех клетках, где активно протекают процессы фагоцитоза с последующим перевариванием захваченного материала (например, в нейтрофильных гранулоцитах, макрофагах, остеокластах).
Лизосомы подразделяются на первичные (неактивные)
ивторичные (активные).
Первичные лизосомы
(гидролазные пузырьки) – округлые пузырьки небольшого размера (обычно около 50 нм диаметром), с мелкозернистым, гомогенным, плотным матриксом. Надежная идентификация первичных лизосом возможна только при гистохимическом выявлении характерных ферментов (кислая фосфатаза
). Первичные лизосомы – неактивные структуры, еще не вступившие в процессы расщепления субстратов.
Вторичные лизосомы
– органеллы, активно участвующие в процессах внутриклеточного переваривания. Диаметр вторичных лизосом обычно составляет 0.5-2 мкм, их форма и структура могут существенно варьировать в зависимости от перевариваемого субстрата, но обычно содержимое вторичных лизосом гетерогенно.
Вторичная лизосома – результат слияния первичной лизосомы с фагосомой или аутофагосомой (Рис.8).
Фаголизосома
формируется путем слияния первичной лизосомы с фагосомой - мембранным пузырьком, содержащим материал, захваченный клеткой извне. Процесс разрушения этого материала называется гетерофагией
. Гетерофагия играет важную роль в функции всех клеток. Особое значение гетерофагия имеет для клеток, осуществляющих защитную функцию, таких как макрофаги и нейтрофильные лейкоциты, которые захватывают и переваривают болезнетворные микроорганизмы.
Аутофаголизосома
образуется при слиянии первичной лизосомы с аутофагосомой - мембранным пузырьком, содержащим собственные компоненты клетки, которые подлежат разрушению. Процесс переваривания внутриклеточного материала называется аутофагией
. Аутофагия обеспечивает постоянное обновление клеточных структур благодаря перевариванию митохондрий, полисом, фрагментов мембран.

Рис.8.
Остаточные тельца
– лизосомы, содержащие непереваренный материал, которые могут находиться в цитоплазме длительное время. В некоторых долгоживущих клетках (нейроны, кардиомиоциты, гепатоциты) в остаточных тельцах накапливается коричневый эндогенный пигмент липофусцин – «пигмент старения».
Дефицит лизосомальных ферментов
может приводить к развитию ряда заболеваний (болезни накопления), вызванных накоплением в клетках непереваренных веществ, которые нарушают функцию клеток. Примерами могут служить: болезнь Хюрлера
, при которой из-за отсутствия α-L-идуронидазы фибробласты и остеобласты накапливают дерматан сульфат, а у больных отмечаются множественные дефекты хондро- и остеогенеза и умственное отставание; болезнь Тэя-Сакса
(из-за недостаточности гексозаминидазы А происходит накопление гликолипидов в нервных клетках и поражается нервная система); болезнь Гоше
(вследствие наследственного дефекта глюкоцереброзидазы гликолипиды накапливаются в макрофагах и поражаются печень и селезенка) и другие.
Пероксисомы
– сферические мембранные органеллы диаметром 0.05 – 1.5 мкм, с умеренно плотным гомогенным или мелкозернистым матриксом. Мелкие пероксисомы встречаются во всех клетках, а крупные пероксисомы – в гепатоцитах, макрофагах, в клетках канальцев почки. Матрикс пероксисом содержит до 50 различных ферментов, важнейшие из которых: каталаза
(маркёр пероксисом), пероксидаза, оксидазы аминокислот, уратоксидаза.
У некоторых видов животных в пероксисомах выявляется более плотная кристаллическая сердцевина – нуклеоид
, состоящая из уратоксидазы. В пероксисомах клеток человека нуклеотида нет, поскольку отсутствует способность метаболизировать ураты.
Функции пероксисом:
Окисление аминокислот и других субстратов;
Защита клетки от действия перекиси водорода, сильного окислителя, образующегося в результате окисления органических соединений, и оказывающего повреждающий эффект на клетку. При этом каталаза пероксисом разлагает перекись водорода на воду и кислород.
Участие в расщеплении жирных кислот;
Участие в обезвреживании ряда веществ (спирт и др.).
Нарушения активности пероксисом вызывает ряд наследственных заболеваний – пероксисомных болезней с тяжелыми нарушениями нервной системы (синдром Целльвегера и др.)

Рис.9.
Цитоскелет
– сложная трехмерная сеть немембранных органелл (рис.9):
· микротрубочек;
· микрофиламентов;
· промежуточных филаментов.
Основная функция цитоскелета – опорно-двигательная:
Поддержание и изменение формы клеток;
Перемещение компонентов внутри клетки;
Транспорт веществ внутрь клетки и из клетки;
Обеспечение подвижности клетки
Микротрубочки
– наиболее крупные компоненты цитоскелета. Микротрубочки – полые цилиндрические образования различной длины, с диаметром 24-25 нм, с толщиной стенки 5 нм.
Стенка микротрубочки состоит из спирально
расположенных нитей – профиламентов
, образованных димерами из глобулярных белковых молекул – α- и β-тубулина
.
Стенка микротрубочки образована 13 субъединицами-профиламентами.
Микротрубочки могут располагаться в цитоплазме в виде отдельных элементов, в виде пучков, где они связаны тонкими поперечными мостиками, или могут частично сливаться друг с другом, образуя дуплеты
(в аксонеме ресничек и жгутиков) и триплеты
(в базальном тельце и центриолях.
Микротрубочки представляют собой лабильную систему, в которой сохраняется равновесие между их постоянной сборкой и диссоциацией.
Центрами организации микротрубочек (ЦОМТ) являются сателлиты – глобулярные белковые структуры, содержащиеся в базальных тельцах ресничек и клеточном центре, а также центромеры хромосом.
Функции микротрубочек:
· поддержание стабильной формы клеток, и порядка распределения её компонентов;
· обеспечение внутриклеточного транспорта, в том числе органелл, пузырьков, секреторных гранул (благодаря некоторым белкам, ассоциированным с микротрубочками);
· образование основы центриолей и ахроматинового веретена деления и обеспечение движения хромосом в процессе митоза;
· образование основы ресничек и жгутиков, а также обеспечение их движения.
Угнетение самосборки микротрубочек при действии на клетку блокаторов (колхицин и др.) вызывает гибель быстроделящихся клеток вследствие отсутствия митотического веретена деления, нарушения транспортных процессов в клетке (аксонный транспорт в нейронах, секреция), изменения форм клетки, дезорганизацию клеточных органелл (в частности, цистерн ЭПС).
Клеточный центр
образован двумя полыми цилиндрическими структурами - центриолями
, которые расположены под прямым углом друг к другу.
Каждая центриоль представляет собой короткий цилиндр длиной ~ 0,5 мкм и диаметром ~ 0,2 мкм, состоящий из 9 триплетов частично слившихся трубочек (А, В и С), связанных поперечными белковыми мостиками (рис.10).

Формула строения центриоли описывается как (9 × 3) + 0
,
так как в центральной части микротрубочки отсутствуют. Каждый триплет центриоли связан с глобулярными белковыми тельцами – сателлитами, от которых отходят микротрубочки, образующие центросферу.
Рис.10.
В неделящейся клетке выявляется одна пара центриолей – диплосома,
которая располагается обычно вблизи ядра. Перед делением клетки в S-периоде интерфазы происходит дупликация центриолей
: под прямым углом к каждой зрелой (материнской) центриоли пары образуется новая (дочерняя) центриоль.
В ранней профазе митоза пары центриолей расходятся к полюсам клетки и служат центрами образования микротрубочек ахроматинового веретена деления.

Реснички
и жгутики
являются выростами цитоплазмы, обладающие подвижностью. Основу ресничек и жгутиков составляет каркас из микротрубочек, называемый аксонемой (рис.11).
Длина ресничек равна 2-10 мкм, а их количество на поверхности одной клетки может составлять до нескольких сотен.
В организме человека жгутик есть только в одном типе клеток – сперматозоидах. При этом один сперматозоид имеет один жгутик длиной 50-70 мкм.
Рис.11.
Аксонема
образована 9периферическими парами микротрубочек (микротрубочки А и В) и одной центрально расположенной парой; такое строение описывается формулой (9 × 2) + 2.
Центральная пара микротрубочек окружена центральной оболочкой, от которой к периферическим дуплетам расходятся радиальные спицы. Периферические дублеты связаны друг с другом мостиками белка нексина, а от микротрубочки А к микротрубочке В соседнего дублета отходят “ручки” из белка динеина,
который обладает АТФ-азной активностью, что необходимо для скольжения соседних дублетов в аксонеме, вызывающих движение (биение) ресничек и жгутиков
Мутации, вызывающих изменения белков ресничек и жгутиков, ведут к различным нарушениям функций клеток. Так, при отсутствии динеиновых ручек (синдром неподвижных ресничек
, или синдром Картагенера
), больные страдают хроническими заболеваниями дыхательной системы и бесплодием (вследствие неподвижности спермиев и нарушений продвижения яйцеклеток по яйцеводу).
В основании каждой реснички или жгутика лежит базальное тельце
, сходное по строению с центриолью. На уровне апикального конца базального тельца микротрубочка С триплета заканчивается, тогда как микротрубочки А и В продолжаются в соответствующие микротрубочки аксонемы реснички. При развитии ресничек или жгутика базальное тельце играет роль матрицы, на которой происходит сборка компонентов аксонемы
Микрофиламенты
– тонкие белковые нити диаметром 5-7 нм, расположенные в цитоплазме поодиночке, в виде сетей или упорядоченными пучками (в скелетной и сердечной мышцах). Основной белок микрофиламентов – актин
– встречается в клетках как в мономерной форме (глобулярный G-актин), так и в виде полимерного фибриллярного F-актина.
Функции микрофиламентов:
В мышечных волокнах и клетках актиновые микрофиламенты образуют упорядоченные пучки и при взаимодействии с миозиновыми филаментами обеспечивают их сокращение.
В немышечных клетках микрофиламенты образуют кортикальную (терминальную) сеть, в которой микрофиламенты сшиты с помощью особых белков (филамин и др.). Кортикальная сеть, с одной стороны, обеспечивает поддержание формы клетки, а с другой - способствует изменениям формы плазмолеммы, обеспечивая, таким образом, функции эндо- и экзоцитоза, миграции клеток, образования псевдоподий.
Микрофиламенты тесно связаны с органеллами, транспортными пузырьками, секреторными гранулами и играют важную роль в их перемещении внутри цитоплазмы.
Микрофиламенты формируют сократимую перетяжку (срединное тельце) при цитотомии, завершающей клеточное деление.
Микрофиламенты участвуют в организации структуры межклеточных соединений (zonula adherens – поясок сцепления).
Микрофиламенты являются основой специальных выростов цитоплазмы – микроворсинок и стереоцилий.
Микроворсинки
– пальцевидные выросты цитоплазмы клетки диаметром 0.1 мкм и длиной 1 мкм, основу которых образуют актиновые микрофиламенты (рис.12).
 Микроворсинки обеспечивают многократное увеличение площади поверхности клетки. На апикальной поверхности некоторых клеток, активно участвующих в процессах расщепления и всасывания веществ, имеется до несколько тысяч микроворсинок, образующих в совокупности щёточную каемку
(эпителий тонкой кишки и почечных канальцев).
Микроворсинки обеспечивают многократное увеличение площади поверхности клетки. На апикальной поверхности некоторых клеток, активно участвующих в процессах расщепления и всасывания веществ, имеется до несколько тысяч микроворсинок, образующих в совокупности щёточную каемку
(эпителий тонкой кишки и почечных канальцев).
Рис.12.
Основа каждой микроворсинки – пучок, содержащий около 40 микрофиламентов, расположенных вдоль её длинной оси. Микрофиламенты имеют поперечные сшивки из белков (фимбрин, виллин), и прикреплены к плазмолемме особыми белковыми мостиками (минимиозин). У основания микроворсинки микрофиламенты пучка вплетаются в терминальную сеть
Стереоцилии
– длинные, иногда ветвящиеся микроворсинки, имеющие каркас из микрофиламентов. Они встречаются редко (например, в главных клетках эпителия протока придатка семенника).
Промежуточные филаменты
– прочные и устойчивые белковые нити толщиной около 10 нм (что является промежуточным
значением между толщиной микротрубочек и микрофиламентов). Промежуточные филаменты располагаются в виде трехмерных сетей в различных участках цитоплазмы, окружают ядро, участвуют в образовании межклеточных контактов (десмосом) и поддерживают форму отростков.
Главная функция
промежуточных филаментов – поддерживающая и опорная.
Промежуточные филаменты в клетках различных типов различаются по своей химической природе и молекулярному весу. Выделяют 6 основных классов промежуточных филаментов
Цитокератины –
промежуточные филаменты, характерные для клеток эпителия. Этот класс включает около 20 близких полипептидов (тонофиламентов). Кератиновые филаменты входят в состав десмосом и полудесмосом, участвуют в образовании рогового вещества в эпителии кожи и являются главным компонентом волос и ногтей.
Десмины
– промежуточные филаменты мышечных тканей (за исключением миоцитов сосудов). Десмины играют важную роль в организации миофибрилл в мышечной ткани и обеспечении сократительной функции
Виментины
– филаменты, характерные для различных клеток мезенхимного
происхождения (фибробласты, макрофаги, остеобласты, эндотелий и гладкие миоциты сосудов).
Нейрофиламенты
– промежуточные филаменты нейронов, которые играют важную роль в поддержании формы отростков нервных клеток.
Глиальные клетки
содержат глиальный фибриллярный кислый белок
и встречаются только в клетках нейроглии (астроциты, олигодендроциты).
Идентификация классов промежуточных филаментов (методами иммуноцитохимии с антителами к данному типу промежуточных филаментов) имеет большое значение в диагностике опухолей, и, следовательно, в прогнозе и выборе противоопухолевого лечения. Так, выявление различных форм кератинов свидетельствует о недифференцированных опухолях эпителиального происхождения, карциномах, аденокарциномах. Десмин является маркёром опухолей мышечного происхождения, а глиальный фибриллярный кислый белок – маркёр опухолей глиального происхождения.
ВКЛЮЧЕНИЯ
В отличие от органелл, включения цитоплазмы – непостоянные компоненты цитоплазмы, возникающие и исчезающие в зависимости от метаболического состояния клеток.
Включения подразделяются на трофические, секреторные, экскреторные и пигментные.
Трофические включения разделяются в зависимости от природы накапливаемого вещества на липидные, углеводные и белковые. Липидные включения – это капли нейтрального жира различного диаметра, которые накапливаются в цитоплазме и служат резервом энергетических субстратов, используемых клеткой. Из углеводных включений наиболее распространены гранулы гликогена (полимер глюкозы), эти включения также используются в качестве источника энергии. Примером белковых включений могут служить запасы белка вителлина в яйцеклетках животных. Они являются источником питания на ранних стадиях развития зародыша.
Секреторные включения имеют вид пузырьков, окруженные мембраной и содержащие биологически активные вещества, которые синтезируются в самой клетке, а затем выделяются (секретируются) во внешнюю среду. К таким включениям относятся секреторные гранулы, содержащие пищеварительные проферменты (зимогеновые гранулы), гормоны, медиаторы и др.
Экскреторные включения по своему строению сходны с секреторными, но в отличие от них, содержат вредные продукты метаболизма, подлежащие удалению из цитоплазмы клеток.
Пигментные включения представляют собой скопления эндогенных (синтезированных клеткой), или экзогенных (захваченных клеткой извне) окрашенных веществ - пигментов. Наиболее распространенными эндогенными пигментами являются гемоглобин, гемосидерин, билирубин, меланин, липофусцин; к экзогенным пигментам относят каротин, различные красители, пылевые частицы и др. Меланин – тёмно-коричневый пигмент, встречающийся в норме в коже, волосах, пигментной оболочке сетчатки в виде меланосом - гранул, окруженных мембраной. Липофусцин – гранулы жёлто-коричневого пигмента из продуктов лизосомного переваривания – накапливается в долгоживущих клетках (нейроны, кардиомиоциты), и поэтому его рассматривают как «пигмент старения».