Mitohondriji se sastoje od. Mitohondriji. Struktura i funkcije membranskih proteinskih kompleksa
Mitohondriji su jedna od najvažnijih komponenti svake stanice. Također se nazivaju hondriosomi. To su zrnasti ili končasti organeli koji su dio citoplazme biljaka i životinja. Oni su proizvođači ATP molekula, koje su toliko potrebne za mnoge procese u stanici.
Što su mitohondriji?
Mitohondriji su energetska baza stanica; njihova se aktivnost temelji na oksidaciji i korištenju energije koja se oslobađa tijekom razgradnje molekula ATP-a. Biolozi na jednostavnim jezikom naziva se stanica za proizvodnju energije za stanice.
Godine 1850. mitohondriji su identificirani kao granule u mišićima. Njihov broj je varirao ovisno o uvjetima rasta: više se akumuliraju u onim stanicama u kojima postoji veliki nedostatak kisika. To se najčešće događa tijekom tjelesne aktivnosti. U takvim tkivima javlja se akutni nedostatak energije, koji se nadopunjuje mitohondrijima.
Pojava pojma i mjesto u teoriji simbiogeneze

Godine 1897., Bend je prvi uveo koncept "mitohondrija" za označavanje granularne i filamentne strukture u kojoj se razlikuju u obliku i veličini: debljina je 0,6 µm, duljina - od 1 do 11 µm. U rijetkim situacijama mitohondriji mogu biti veliki i razgranati.
Teorija simbiogeneze daje jasnu ideju o tome što su mitohondriji i kako su se pojavili u stanicama. Kaže da je hondriosom nastao u procesu oštećenja bakterijskih stanica, prokariota. Budući da nisu mogli samostalno koristiti kisik za stvaranje energije, to ih je spriječilo u potpunom razvoju, dok su se progenoti mogli nesmetano razvijati. Tijekom evolucije, veza između njih omogućila je progenotima da prenesu svoje gene na eukariote. Zahvaljujući tom napretku, mitohondriji više nisu neovisni organizmi. Njihov genski fond ne može se u potpunosti realizirati, jer je djelomično blokiran enzimima koji su prisutni u bilo kojoj stanici.
Gdje oni žive?
Mitohondriji su koncentrirani u onim područjima citoplazme gdje se javlja potreba za ATP-om. Na primjer, u mišićnom tkivu srca nalaze se u blizini miofibrila, au spermatozoidima tvore zaštitnu kamuflažu oko osi vrpce. Tamo generiraju puno energije kako bi se "rep" vrtio. Tako se spermij kreće prema jajnoj stanici.
U stanicama novi mitohondriji nastaju jednostavnom diobom prethodnih organela. Tijekom njega se čuvaju sve nasljedne informacije.
Mitohondriji: kako izgledaju
Oblik mitohondrija podsjeća na cilindar. Često se nalaze u eukariota, zauzimajući od 10 do 21% volumena stanice. Njihove veličine i oblici jako variraju i mogu se mijenjati ovisno o uvjetima, ali je širina konstantna: 0,5-1 mikrona. Kretanje hondriosoma ovisi o mjestima u stanici gdje se energija brzo rasipa. Kreću se kroz citoplazmu koristeći za kretanje strukture citoskeleta.

Zamjena za mitohondrije različitih veličina, koji rade odvojeno jedni od drugih i opskrbljuju energijom određene zone citoplazme, su dugi i razgranati mitohondriji. Oni su u stanju opskrbljivati energijom područja stanica koja se nalaze daleko jedna od druge. Sličan suradnja Hondriosomi se uočavaju ne samo u jednostaničnim organizmima, već iu višestaničnim. Najviše složena struktura Hondriosomi se nalaze u skeletnim mišićima sisavaca, gdje se najveći razgranati hondriosomi spajaju jedan s drugim pomoću intermitohondrijskih kontakata (IMC).
To su uski razmaci između susjednih mitohondrijskih membrana. Ovaj prostor ima visoku gustoću elektrona. MMK su češći u stanicama gdje se vežu zajedno s radnim hondriosomima.
Da bismo bolje razumjeli problem, potrebno je ukratko opisati značaj mitohondrija, strukturu i funkcije ovih nevjerojatnih organela.
Kako su izgrađeni?
Da biste razumjeli što su mitohondriji, morate znati njihovu strukturu. Ovaj neobičan izvor energije sfernog je oblika, ali često izdužen. Dvije membrane nalaze se blizu jedna druge:
- vanjski (glatki);
- unutarnji, koji tvori lisnate (cristae) i cjevaste (tubule) izdanke.
Osim veličine i oblika mitohondrija, njihova građa i funkcija su jednaki. Hondriosom je omeđen dvjema membranama veličine 6 nm. Vanjska membrana mitohondrija nalikuje spremniku koji ih štiti od hijaloplazme. Unutarnja membrana je odvojena od vanjske membrane područjem širine 11-19 nm. Posebnost unutarnje membrane je njezina sposobnost da strši u mitohondrije, poprimajući oblik spljoštenih grebena.

Unutarnja šupljina mitohondrija ispunjena je matriksom fino zrnate strukture, gdje se ponekad nalaze niti i granule (15-20 nm). Matrične niti stvaraju organele i granule male veličine- ribosomi, mitohondriji.
U prvoj fazi odvija se u hijaloplazmi. U ovoj fazi dolazi do početne oksidacije supstrata ili glukoze do Ovi se postupci odvijaju bez kisika – anaerobna oksidacija. Sljedeći stupanj proizvodnje energije sastoji se od aerobne oksidacije i razgradnje ATP-a, taj se proces odvija u mitohondrijima stanica.
Što rade mitohondriji?
Glavne funkcije ove organele su:

Prisutnost vlastite deoksiribonukleinske kiseline u mitohondrijima još jednom potvrđuje simbiotičku teoriju o pojavi ovih organela. Također, uz svoj glavni posao, uključeni su u sintezu hormona i aminokiselina.
Patologija mitohondrija
Mutacije koje se javljaju u mitohondrijskom genomu dovode do depresivnih posljedica. Ljudski nositelj je DNK, koja se na potomke prenosi s roditelja, dok se mitohondrijski genom prenosi samo s majke. Ova činjenica se objašnjava vrlo jednostavno: djeca dobivaju citoplazmu s hondriosomima koji su u njoj zatvoreni zajedno s ženskim jajetom; oni su odsutni u spermi. Žene s ovim poremećajem mogu prenijeti mitohondrijske bolesti na svoje potomke, ali bolesni muškarac ne može.

U normalnim uvjetima Hondriosomi imaju istu kopiju DNA – homoplazmu. U genomu mitohondrija mogu se pojaviti mutacije, a heteroplazmija nastaje zbog suživota zdravih i mutiranih stanica.
Zahvaljujući suvremenoj medicini danas je identificirano više od 200 bolesti čiji je uzrok bila mutacija mitohondrijske DNA. Ne u svim slučajevima, ali mitohondrijske bolesti dobro reagiraju na terapijsko održavanje i liječenje.
Tako smo shvatili pitanje što su mitohondriji. Kao i svi drugi organeli, vrlo su važni za stanicu. Neizravno sudjeluju u svim procesima koji zahtijevaju energiju.
Geni koji su tijekom evolucije ostali u “energetskim stanicama stanice” pomažu u izbjegavanju problema upravljanja: ako se nešto pokvari u mitohondriju, on to može sam popraviti, ne čekajući dopuštenje iz “centra”.
Naše stanice dobivaju energiju uz pomoć posebnih organela zvanih mitohondriji, koji se često nazivaju energetskim stanicama stanice. Izvana izgledaju kao spremnici s dvostrukom stijenkom, a unutarnja stijenka je vrlo neravna, s brojnim jakim udubljenjima.
Stanica s jezgrom (plavo) i mitohondrijima (crveno). (Fotografija NICHD/Flickr.com)
Mitohondriji u presjeku, izdanci unutarnje membrane vidljivi su kao uzdužne unutarnje pruge. (Fotografija Visuals Unlimited/Corbis.)
U mitohondrijima se događa ogroman broj biokemijskih reakcija, tijekom kojih se molekule “hrane” postupno oksidiraju i razgrađuju, a njihova energija kemijske veze pohranjeni u obliku pogodnom za stanicu. No, osim toga, te "energetske stanice" imaju vlastitu DNK s genima, koju opslužuju vlastiti molekularni strojevi koji osiguravaju sintezu RNK, a zatim sintezu proteina.
Vjeruje se da su mitohondriji u davnoj prošlosti bili samostalne bakterije kojima su se hranili neki drugi jednostanični bići (najvjerojatnije arheje). Ali jednog dana "grabežljivci" su iznenada prestali probavljati progutane protomitohondrije, zadržavajući ih u sebi. Započelo je dugo trljanje simbionta jednog o drugog; kao rezultat toga, one koje su progutane znatno su pojednostavile svoju strukturu i postale su unutarstanične organele, a njihovi „vlasnici“ su se zahvaljujući učinkovitijoj energiji mogli dalje razvijati, sve više i više složenih oblikaživota, sve do biljaka i životinja.
Da su mitohondriji nekada bili neovisni svjedoče ostaci njihovog genetskog aparata. Naravno, ako živite unutra sa svime gotovim, nestaje potreba za sadržavanjem vlastitih gena: DNK modernih mitohondrija u ljudskim stanicama sadrži samo 37 gena - nasuprot 20-25 tisuća onih sadržanih u nuklearnoj DNK. Tijekom milijuna godina evolucije, mnogi od mitohondrijskih gena preselili su se u jezgru stanice: proteini koje kodiraju sintetiziraju se u citoplazmi i zatim transportiraju u mitohondrije. Međutim, odmah se postavlja pitanje zašto je 37 gena ipak ostalo tamo gdje je bilo?
Mitohondriji su, ponavljamo, prisutni u svim eukariotskim organizmima, odnosno u životinjama, biljkama, gljivama i protozoama. Ian Johnston ( Iain Johnston) sa Sveučilišta u Birminghamu i Ben Williams ( Ben P. Williams) s Instituta Whitehead analizirali su više od 2000 mitohondrijskih genoma uzetih iz različitih eukariota. Pomoću posebnog matematičkog modela znanstvenici su uspjeli shvatiti koji će geni vjerojatnije ostati u mitohondrijima tijekom evolucije.
Vanjska membrana
Unutarnja membrana
Matrica m-na, matrica, cristas. ima glatke konture i ne stvara udubljenja ili nabore. Zauzima oko 7% ukupne površine stanične membrane. Debljina joj je oko 7 nm, nije vezana ni za jednu drugu membranu citoplazme i zatvorena je u sebe, tako da je membranska vrećica. Odvaja vanjsku membranu od unutarnje intermembranski prostorširok oko 10-20 nm. Unutarnja membrana (debljine oko 7 nm) ograničava stvarni unutarnji sadržaj mitohondrija,
njegov matriks ili mitoplazma. Karakteristična značajka Unutarnja membrana mitohondrija je njihova sposobnost stvaranja brojnih invaginacija u mitohondrije. Takve invaginacije najčešće imaju oblik ravnih grebena ili krista. Razmak između membrana u kristi je oko 10-20 nm. Često se kriste mogu granati ili formirati prstaste izdanke, savijati se i nemaju jasnu orijentaciju. U najjednostavnijih, jednostaničnih algi, te u nekim stanicama viših biljaka i životinja, izdanci unutarnje membrane imaju oblik cjevčica (cjevaste kriste).
Mitohondrijski matriks ima fino zrnatu homogenu strukturu; ponekad se u njemu otkrivaju tanki filamenti skupljeni u kuglu (oko 2-3 nm) i granule oko 15-20 nm. Sada je postalo poznato da su filamenti mitohondrijskog matriksa DNA molekule unutar mitohondrijskog nukleoida, i male granule– mitohondrijski ribosomi.
Funkcije mitohondrija
1. Sinteza ATP-a odvija se u mitohondrijima (vidi Oksidativna fosforilacija)
PH međumembranskog prostora ~4, pH matriksa ~8 | sadržaj proteina u m: 67% - matrica, 21% - vanjski m-on, 6% - unutarnji m-on i 6% - u intersticijskoj masi
Handrioma– jedinstveni mitohondrijski sustav
vanjska m-na: porini-pore dopuštaju prolaz do 5 kD | unutarnja m-na: kardiolipin – čini m-n nepropusnom za ione |
povremena proizvodnja: skupine enzima fosforiliraju nukleotide i šećere nukleotida
unutarnja m-na:
matriks: metabolički enzimi - oksidacija lipida, oksidacija ugljikohidrata, ciklus trikarboksilne kiseline, Krebsov ciklus
Podrijetlo od bakterija: ameba Pelomyxa palustris ne sadrži eukariote, živi u simbiozi s aerobnim bakterijama | vlastiti DNK | procesi slični bakterijama
Mitohondrijska DNA
Miohondrijska podjela
replicirano
u međufazi | replikacija nije povezana sa S-fazom | tijekom CL ciklusa, mitohi se dijele jednom na dva, tvoreći suženje, suženje prvo na unutarnjoj strani | ~16,5 kb | kružni, kodira 2 rRNA, 22 tRNA i 13 proteina |
transport proteina: signalni peptid | amfifilni uvojak | mitohondrijski receptor za prepoznavanje |
Oksidativne fosforilacije
Transportni lanac elektrona
ATP sintaza
u stanici jetre, m živi ~20 dana, dioba mitohondrija stvaranjem suženja
16569 bp = 13 proteina, 22 tRNA, 2 pRNA | glatka vanjska membrana (porini - propusnost proteina do 10 kDa) presavijena unutarnja membrana (kriste) matrica (75% proteina: proteini prijenosnici, proteini, komponente dišnog lanca i ATP sintaze, kardiolipin) matrica ( obogaćena tvarima citrata ciklus) isprekidana proizvodnja 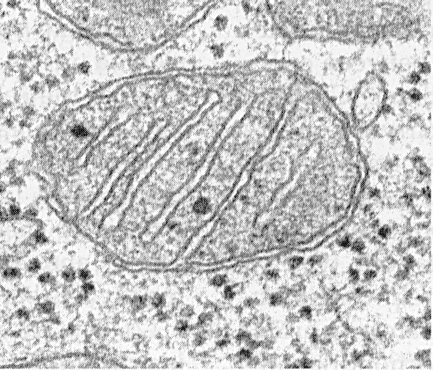
Podrijetlo mitohondrija
Intermembranski prostor
Intermembranski prostor je prostor između vanjske i unutarnje membrane mitohondrija. Debljina mu je 10-20 nm. Budući da je vanjska membrana mitohondrija propusna za male molekule i ione, njihova se koncentracija u periplazmatskom prostoru malo razlikuje od one u citoplazmi. Naprotiv, veliki proteini zahtijevaju specifične signalne peptide za transport iz citoplazme u periplazmatski prostor; stoga su proteinske komponente periplazmatskog prostora i citoplazme različite. Jedan od proteina sadržanih u periplazmatskom prostoru je citokrom c, jedna od komponenti dišnog lanca mitohondrija.
Unutarnja membrana
Unutarnja membrana tvori brojne češljaste nabore - kriste, koje znatno povećavaju njezinu površinu i, primjerice, u stanicama jetre čini oko trećinu svih staničnih membrana. Karakteristična značajka sastava unutarnje mitohondrijske membrane je prisutnost kardiolipina u njoj - posebnog fosfolipida koji sadrži četiri masne kiseline i čini membranu apsolutno nepropusnom za protone. Još jedna značajka unutarnje mitohondrijske membrane je vrlo visok sadržaj proteina (do 70% težine), predstavljen transportnim proteinima, enzimima respiratornog lanca i velikim kompleksima ATP sintetaze. Unutarnja membrana mitohondrija, za razliku od vanjske, nema posebne otvore za transport malih molekula i iona; na njemu, na strani okrenutoj prema matriksu, nalaze se posebne molekule ATP sintaze, koje se sastoje od glavice, peteljke i baze. Kada protoni prolaze kroz njih, dolazi do sinteze ATP-a. U podnožju čestica, ispunjavajući cijelu debljinu membrane, nalaze se komponente dišnog lanca. Vanjska i unutarnja membrana se na nekim mjestima dodiruju; postoji poseban proteinski receptor koji potiče transport mitohondrijskih proteina kodiranih u jezgri u mitohondrijski matriks.
Matrica
Matrica je prostor ograničen unutarnjom membranom. Matrica (ružičasta tvar) mitohondrija sadrži enzimske sustave za oksidaciju piruvata, masnih kiselina, kao i enzime ciklusa trikarboksilnih kiselina (Krebsov ciklus). Osim toga, ovdje se nalaze i mitohondrijska DNA, RNA i mitohondrijski vlastiti aparat za sintezu proteina.
Mitohondrijska DNA
Mitohondrijska DNA smještena u matrici je zatvorena kružna dvolančana molekula, u ljudskim stanicama veličine 16569 parova nukleotida, što je otprilike 10 5 puta manje od DNA lokalizirane u jezgri. Ukupno, mitohondrijska DNA kodira 2 rRNA, 22 tRNA i 13 podjedinica enzima respiratornog lanca, što ne čini više od polovice proteina koji se nalaze u njoj. Konkretno, sedam podjedinica ATP sintetaze, tri podjedinice citokrom oksidaze i jedna podjedinica ubikinol-citokroma kodirane su pod kontrolom mitohondrijskog genoma. S- reduktaza. U ovom slučaju, svi proteini osim jedne, dvije ribosomske i šest tRNA prepisuju se s težeg (vanjskog) lanca DNA, a 14 drugih tRNA i jedan protein prepisuju se iz lakšeg (unutarnjeg) lanca.
Usprkos ovoj pozadini, biljni mitohondrijski genom mnogo je veći i može doseći 370 000 parova nukleotida, što je otprilike 20 puta više od gore opisanog ljudskog mitohondrijalnog genoma. Broj gena ovdje je također približno 7 puta veći, što je popraćeno pojavom u biljnim mitohondrijima dodatnih putova prijenosa elektrona koji nisu povezani sa sintezom ATP-a.
Dakle, cjelokupna reakcija koju kataliziraju enzimi dišnog lanca je oksidacija NADH s kisikom u vodu. U biti, ovaj se proces sastoji od postupnog prijenosa elektrona između atoma metala prisutnih u prostetskim skupinama proteinskih kompleksa dišnog lanca, pri čemu svaki sljedeći kompleks ima veći afinitet prema elektronu od prethodnog. U tom se slučaju sami elektroni prenose duž lanca dok se ne spoje s molekulskim kisikom, koji ima najveći afinitet prema elektronima. Oslobođena energija u ovom slučaju pohranjuje se u obliku elektrokemijskog (protonskog) gradijenta s obje strane unutarnje mitohondrijske membrane. Vjeruje se da se tijekom transporta elektronskih parova kroz dišni lanac pumpa od tri do šest protona.
Završna faza funkcioniranja mitohondrija je stvaranje ATP-a, koje provodi poseban makromolekularni kompleks molekulske mase 500 kDa ugrađen u unutarnju membranu. Ovaj kompleks, nazvan ATP sintetaza, katalizira sintezu ATP-a pretvaranjem energije transmembranskog elektrokemijskog gradijenta vodikovih protona u energiju visokoenergetske veze molekule ATP-a.
sinteza ATP-a
U strukturnom i funkcionalnom smislu, ATP sintaza se sastoji od dva velika fragmenta, označena simbolima F 1 i F 0. Prvi od njih (faktor spajanja F1) okrenut je prema mitohondrijskom matriksu i primjetno strši iz membrane u obliku kuglaste formacije visine 8 nm i širine 10 nm. Sastoji se od devet podjedinica koje predstavljaju pet vrsta proteina. Polipeptidni lanci od tri α podjedinice i isto toliko β podjedinica raspoređeni su u proteinske globule slične strukture, koje zajedno tvore heksamer (αβ) 3, koji izgleda kao blago spljoštena lopta. Poput tijesno zbijenih kriški naranče, uzastopne α i β podjedinice tvore strukturu koju karakterizira os simetrije trećeg reda s kutom rotacije od 120°. U središtu ovog heksamera nalazi se γ podjedinica, koju čine dva produžena polipeptidna lanca i nalikuje blago deformiranom zakrivljenom štapiću dugom oko 9 nm. pri čemu Donji dioγ podjedinica strši iz kuglice za 3 nm prema membranskom kompleksu F 0 . Također unutar heksamera nalazi se manja ε podjedinica povezana s γ. Posljednja (deveta) podjedinica označena je simbolom δ i nalazi se s vanjske strane F 1 .
Membranski dio ATP sintaze, nazvan faktor spajanja F0, je hidrofobni proteinski kompleks koji prodire kroz membranu i ima dva polukanala unutra za prolaz protona vodika. Ukupno F 0 kompleks uključuje jednu proteinsku podjedinicu tipa A, dva primjerka podjedinice b, kao i 9 do 12 kopija male podjedinice c. Podjedinica A (molekularna masa 20 kDa) potpuno je uronjen u membranu, gdje formira šest α-spiralnih odsječaka koji je prelaze. Podjedinica b(molekularne težine 30 kDa) sadrži samo jednu relativno kratku α-spiralnu regiju uronjenu u membranu, a ostatak zamjetno strši iz membrane prema F 1 i vezan je za δ podjedinicu koja se nalazi na njezinoj površini. Svaka od 9-12 kopija podjedinice c(molekulske težine 6-11 kDa) je relativno mali protein od dvije hidrofobne α-spirale međusobno povezane kratkom hidrofilnom petljom usmjerenom prema F 1, a zajedno čine jedinstvenu cjelinu u obliku cilindra uronjenog u membranu. . Podjedinica γ koja strši iz kompleksa F 1 prema F 0 precizno je uronjena u ovaj cilindar i prilično je čvrsto pričvršćena za njega.
Dakle, u molekuli ATP sintaze mogu se razlikovati dvije skupine proteinskih podjedinica koje se mogu usporediti s dva dijela motora: rotor i stator. "Stator" je nepomičan u odnosu na membranu i uključuje sferni heksamer (αβ) 3 koji se nalazi na njegovoj površini i δ podjedinicu, kao i podjedinice a I b membranski kompleks F0. „Rotor“, pokretljiv u odnosu na ovu strukturu, sastoji se od podjedinica γ i ε, koje su, vidljivo stršeći iz kompleksa (αβ) 3, povezane s prstenom podjedinica uronjenih u membranu. c.
Sposobnost sintetiziranja ATP-a svojstvo je jednog kompleksa F 0 F 1, povezanog s prijenosom vodikovih protona kroz F 0 u F 1, u potonjem od kojih se nalaze katalitički centri koji pretvaraju ADP i fosfat u molekulu ATP-a. . Pokretačka snaga za rad ATP sintaze je protonski potencijal stvoren na unutarnjoj mitohondrijskoj membrani kao rezultat rada transportnog lanca elektrona.
Sila koja pokreće "rotor" ATP sintaze javlja se kada razlika potencijala između vanjskog i unutarnje strane membrana > 220 mV, a osigurava se protokom protona koji teče kroz poseban kanal u F0, koji se nalazi na granici između podjedinica a I c. U ovom slučaju put prijenosa protona uključuje sljedeće strukturne elemente:
- Dva nekoaksijalno smještena "polukanala", od kojih prvi osigurava dovod protona iz međumembranskog prostora do bitnih funkcionalnih skupina F0, a drugi osigurava njihov izlazak u matriks mitohondrija;
- Prsten podjedinica c, od kojih svaki u svom središnjem dijelu sadrži protoniranu karboksilnu skupinu, sposobnu vezati H + iz intermembranskog prostora i otpustiti ih kroz odgovarajuće protonske kanale. Kao rezultat periodičnih pomaka podjedinica S, uzrokovan protokom protona kroz protonski kanal, γ podjedinica rotira, uronjena u prsten podjedinica S.
Dakle, katalitička aktivnost ATP sintaze izravno je povezana s rotacijom njezina "rotora", u kojem rotacija γ podjedinice uzrokuje istovremenu promjenu konformacije sve tri katalitičke podjedinice β, što u konačnici osigurava funkcioniranje enzima . U ovom slučaju, u slučaju stvaranja ATP-a, "rotor" se okreće u smjeru kazaljke na satu brzinom od četiri okretaja u sekundi, a sama rotacija se događa u diskretnim skokovima od 120°, od kojih svaki prati stvaranje jedne molekule ATP-a. .
Izravna funkcija sinteze ATP-a je lokalizirana na β-podjedinicama F1 konjugacijskog kompleksa. U ovom slučaju, prvi čin u lancu događaja koji dovodi do stvaranja ATP-a je vezanje ADP-a i fosfata na aktivno središte slobodna β-podjedinica koja se nalazi u stanju 1. Zbog energije vanjskog izvora (protonska struja) dolazi do konformacijskih promjena u kompleksu F 1, uslijed čega se ADP i fosfat čvrsto vežu za katalitički centar (stanje 2) , gdje između njih postaje moguća tvorba kovalentne veze, što dovodi do stvaranja ATP-a. U ovoj fazi ATP sintaze, enzim ne zahtijeva praktički nikakvu energiju, koja će biti potrebna u sljedećoj fazi za oslobađanje čvrsto vezane molekule ATP iz enzimatskog središta. Stoga je sljedeća faza djelovanja enzima da, kao rezultat energetski ovisne strukturne promjene u kompleksu F 1, katalitička β-podjedinica koja sadrži čvrsto vezanu molekulu ATP prelazi u stanje 3, u kojem veza ATP-a pri čemu je katalitički centar oslabljen. Kao rezultat toga, molekula ATP-a napušta enzim, a β-podjedinica se vraća u svoje prvobitno stanje 1, čime se osigurava kruženje enzima.
Rad ATP sintaze povezan je s mehaničkim pokretima njezinih pojedinih dijelova, što omogućuje klasificiranje ovog procesa kao posebne vrste fenomena koji se naziva "rotacijska kataliza". Slično struja u namotu elektromotora pomiče rotor u odnosu na stator, usmjereni prijenos protona kroz ATP sintetazu uzrokuje rotaciju pojedinih podjedinica faktora konjugacije F 1 u odnosu na druge podjedinice enzimskog kompleksa, kao rezultat koji ovaj jedinstveni uređaj za stvaranje energije obavlja kemijski rad – sintetizira molekule ATP-a. Nakon toga, ATP ulazi u citoplazmu stanice, gdje se troši na razne procese ovisne o energiji. Takav prijenos provodi poseban enzim ATP/ADP translokaza ugrađen u membranu mitohondrija, koji izmjenjuje novosintetizirani ATP za citoplazmatski ADP, što jamči sigurnost pula adenil nukleotida unutar mitohondrija.
Rječnik sinonima
Mitohondriji. Vidi plastosom. (
Mitohondriji
Mitohondriji su štapićaste ili ovalne strukture (grč. mitos- nit, hondros- granula). Nalaze se u svim životinjskim stanicama (isključujući zrele crvene krvne stanice): u višim biljkama, algama i protozoama. Nema ih samo kod prokariotskih bakterija.
Ove organele prvi je otkrio i opisao krajem prošlog stoljeća Altman. Nešto kasnije te su strukture nazvane mitohondrijima. Hogeboom je 1948. ukazao na važnost mitohondrija kao središta staničnog disanja, a 1949. Kennedy i Lehninger ustanovili su da se u mitohondrijima odvija ciklus oksidativne fosforilacije. Tako je dokazano da mitohondriji služe kao mjesto za stvaranje energije.
Mitohondriji su vidljivi u konvencionalnom svjetlosnom mikroskopu pomoću posebnih metoda bojenja. U mikroskopu s faznim kontrastom iu "tamnom polju" mogu se promatrati u živim stanicama.
Struktura, dimenzije, oblik mitohondriji su vrlo varijabilni. To prvenstveno ovisi o funkcionalnom stanju stanica. Na primjer, utvrđeno je da se u motornim neuronima muha koje lete neprekidno 2 sata pojavljuje ogroman broj kuglastih mitohondrija, dok je kod muha sa slijepljenim krilima broj mitohondrija znatno manji i štapićastog su oblika (L. B. Levinson). Po obliku mogu biti nitaste, štapićaste, zaobljene i bučice, čak i unutar iste ćelije.
Mitohondriji su lokalizirani u stanici, u pravilu, ili u onim područjima gdje se troši energija ili u blizini nakupina supstrata (na primjer, kapljica lipida), ako ih ima.
Stroga orijentacija mitohondrija nalazi se duž bičeva spermija, u poprečno-prugastom mišićnom tkivu, gdje se nalaze duž miofibrila, u epitelu bubrežnih tubula lokalizirani su u invaginacijama bazalne membrane itd.
Broj mitohondrija u stanicama ima karakteristike organa, na primjer, stanice jetre štakora sadrže od 100 do 2500 mitohondrija, a stanice sabirnih kanalića bubrega - 300, u spermi različitih životinjskih vrsta od 20 do 72, u golemoj amebi Kaos kaos njihov broj doseže 500 000 mitohondrija veličine od 1 do 10 mikrona.
Ultramikroskopska struktura mitohondrija je ista, bez obzira na njihov oblik i veličinu. Prekriveni su s dvije lipoproteinske membrane: vanjskom i unutarnjom. Između njih nalazi se međumembranski prostor.
Invaginacije unutarnje membrane koje strše u tijelo mitohondrija nazivaju se Kristami. Raspored krista u mitohondrijima može biti poprečan ili uzdužan. Oblik krista može biti jednostavan ili razgranat. Ponekad čine složenu mrežu. U nekim stanicama, npr. u stanicama zone glomeruloze nadbubrežne žlijezde, kriste izgledaju poput cjevčica. Broj krista izravno je proporcionalan intenzitetu oksidativnih procesa koji se odvijaju u mitohondrijima. Na primjer, u mitohondrijima kardiomiocita ima ih nekoliko puta više nego u mitohondrijima hepatocita. Prostor zatvoren unutarnjom membranom čini unutarnju komoru mitohondrija. U njemu se između krista nalazi mitohondrijski matriks – relativno gusta tvar elektrona.
Proteine unutarnje membrane sintetiziraju mitoribosomi, a proteine vanjske membrane sintetiziraju citoribosomi.
"Vanjska membrana mitohondrija je u mnogočemu slična membranama ER. Ima ih malo u membranskom prostoru. Dakle, enzimi Krebsovog ciklusa i oksidacije masnih kiselina koncentrirani su u mitohondrijskom matriksu. U unutarnjem transportnom lancu elektrona, fosforilacijski enzimi (stvaranje ATP-a iz ADP-a) i brojni transportni sustavi su lokalizirani u membrani.
Osim proteina i lipida, sastav mitohondrijskih membrana uključuje RNA i DNA, potonji ima genetsku specifičnost i razlikuje se u svojim fizikalno-kemijskim svojstvima od nuklearne DNA.
Elektronsko mikroskopska istraživanja otkrila su da je površina vanjske membrane prekrivena malim sfernim elementarnim česticama. Unutarnja membrana i kriste sadrže slične elementarne čestice na "nogama", takozvanim tijelima gljiva. Sastoje se od tri dijela: kuglaste glave (promjera 90-100 A°), cilindričnog kraka, dugačkog 5 nm i širokog 3-4 nm, te baze dimenzija 4 x 11 nm. Glave tijela gljive povezane su s fosforilacijom, a potom je otkriveno da glave sadrže enzim s aktivnošću ATP-ida.
U intermembranskom prostoru nalazi se tvar s nižom elektronskom gustoćom od matrice. Omogućuje komunikaciju između membrana i opskrbljuje pomoćne koenzimske katalizatore za enzime smještene u obje membrane.
Danas je poznato da je vanjska membrana mitohondrija vrlo propusna za tvari niske molekularne težine, posebice proteinske spojeve. Unutarnja membrana mitohondrija je selektivno propusna. Praktično je nepropustan za anione (Cl -1, Br -1, SO 4 -2, HCO 3 -1, Sn +2, Mg +2 katione, niz šećera i većinu aminokiselina, dok Ca 2+, Mn 2 +, fosfat , polikarboksilne kiseline lako prodiru kroz nju. Postoje dokazi o prisutnosti nekoliko nosača specifičnih za određene skupine prodornih aniona i kationa koji se odvijaju pomoću energije ATP-aznog sustava ili električni potencijal koji nastaje na membrani kao rezultat rada dišnog lanca čak se i ATP sintetiziran u mitohondrijima može osloboditi pomoću transportera.
Mitohondrijska matrica predstavljena je fino zrnatom elektronskom gustom tvari. Sadrži mitoribosome, fibrilarne strukture koje se sastoje od molekula DNA i granula promjera većeg od 200A ◦ koje čine soli: Ca 3 (PO 4), Ba 3 (PO 4) 2, Mg 3 (PO 4). Vjeruje se da granule služe kao spremnik iona Ca +2 i Mg +2. Njihov broj raste s promjenama propusnosti mitohondrijskih membrana.
Prisutnost DNA u mitohondrijima osigurava sudjelovanje mitohondrija u sintezi RNA i specifičnih proteina, a također ukazuje na postojanje citoplazmatskog nasljeđa. Svaki mitohondrij sadrži, ovisno o veličini, jednu ili više molekula DNA (od 2 do 10). Molekularna težina mitohondrijske DNA je oko (30-40) * 10 6 u protozoama, kvascima i gljivama. Kod viših životinja postoji oko (9-10) * 10 6.
Njegova duljina u kvascu je približno 5 mikrona, u biljkama - 30 mikrona. Količina genetskih informacija sadržanih u mitohondrijskoj DNK je mala: sastoji se od 15-75 tisuća parova baza, koji u prosjeku mogu kodirati 25-125 proteinskih lanaca s molekularnom težinom od oko 40.000.
Mitohondrijska DNK razlikuje se od nuklearne DNK na više načina: više velika brzina sinteze (5-7 puta), otporniji je na djelovanje DNaze, dvoprstenasta je molekula, sadrži više gvanina i citozina, denaturira na višoj temperaturi i lakše se obnavlja. Međutim, mitohondrijski sustav ne sintetizira sve mitohondrijske proteine. Dakle, sinteza citokroma C i drugih enzima osigurava se informacijama sadržanim u jezgri. Vitamini A, B2, B12, K, E, kao i glikogen su lokalizirani u matriksu mitohondrija.
Funkcija mitohondrija sastoji se u stvaranju energije potrebne za život stanica. Kao izvor energije u stanici mogu poslužiti različiti spojevi: bjelančevine, masti, ugljikohidrati. Međutim, jedini supstrat koji se odmah uključuje u energetske procese je glukoza.
Biološki procesi, kao rezultat kojih se stvara energija u mitohondrijima, mogu se podijeliti u 3 skupine: I. skupina - oksidativne reakcije, uključujući dvije faze: anaerobnu (glikoliza) i aerobnu. II skupina - defosforilacija, razgradnja ATP-a i oslobađanje energije. III skupina - fosforilacija povezana s procesom oksidacije.
Proces oksidacije glukoze u početku se odvija bez sudjelovanja kisika (anaerobno ili glikolitički) u pirogrožđanu ili mliječnu kiselinu.
Međutim, oslobađa se samo mala količina energije. Zatim su te kiseline uključene u oksidacijske procese koji se odvijaju uz sudjelovanje kisika, tj. aerobne su. Kao rezultat procesa oksidacije pirogrožđane i mliječne kiseline, nazvanog Krebsov ciklus, ugljični dioksid, voda i veliki broj energije.
Nastala energija se ne oslobađa u obliku topline, što bi dovelo do pregrijavanja stanica i smrti cijelog organizma, već se akumulira u obliku pogodnom za skladištenje i transport u obliku adenozin trifosforne kiseline (ATP). Sinteza ATP-a odvija se iz ADP-a i fosforne kiseline i stoga se naziva fosforilacija.
U zdravim stanicama fosforilacija je povezana s oksidacijom. U bolestima se konjugacija može razdvojiti, pa se supstrat oksidira, ali ne dolazi do fosforilacije, a oksidacija prelazi u toplinu, a sadržaj ATP-a u stanicama opada. Kao rezultat toga, temperatura raste i smanjuje se funkcionalna aktivnost stanica.
Dakle, glavna funkcija mitohondrija je proizvodnja gotovo sve energije stanice i dolazi do sinteze komponenti potrebnih za aktivnost same organele, enzima "respiratornog ansambla", fosfolipida i proteina.
Drugi aspekt aktivnosti mitohondrija je njihovo sudjelovanje u specifičnim sintezama, na primjer, u sintezi steroidnih hormona i pojedinačnih lipida. U jajnim stanicama raznih životinja nastaju nakupine žumanjka u mitohondrijima i one gube svoj osnovni sustav. Potrošeni mitohondriji također mogu akumulirati proizvode izlučivanja.
U nekim slučajevima (jetra, bubrezi) mitohondriji su sposobni akumulirati štetne tvari i otrove koji ulaze u stanicu, izolirati ih od glavne citoplazme i djelomično blokirati štetne učinke tih tvari. Dakle, mitohondriji su sposobni preuzeti funkcije drugih staničnih organela kada je to potrebno za potpuno osiguranje određenog procesa u normalnim uvjetima ili u ekstremnim uvjetima.
Biogeneza mitohondrija. Mitohondriji su obnovljive strukture s prilično kratkim životnim ciklusom (u stanicama jetre štakora, na primjer, poluživot mitohondrija obuhvaća oko 10 dana). Mitohondriji nastaju kao rezultat rasta i diobe prethodnih mitohondrija. Njihova se dioba može dogoditi na tri načina: sužavanjem, pupanjem malih dijelova i pojavom mitohondrija kćeri unutar majke. Diobi (razmnožavanju) mitohondrija prethodi razmnožavanje vlastitog genetskog sustava – mitohondrijske DNA.
Dakle, prema stajalištima većine istraživača, formiranje mitohondrija događa se uglavnom kroz samoreprodukciju de novo.